Органы чувств

Разнообразие организмов: одноклеточные и многоклеточные; автотрофы, гетеротрофы
Одноклеточные и многоклеточные организмы
Необычайное разнообразие живых существ на планете вынуждает находить различные критерии для их классификации. Так, их относят к клеточным и неклеточным формам жизни, поскольку клетки являются единицей строения почти всех известных организмов — растений, животных, грибов и бактерий, тогда как вирусы являются неклеточными формами.
В зависимости от количества клеток, входящих в состав организма, и степени их взаимодействия выделяют одноклеточные, колониальные и многоклеточные организмы. Несмотря на то, что все клетки сходны морфологически и способны осуществлять обычные функции клетки (обмен веществ, поддержание гомеостаза, развитие и др.), клетки одноклеточных организмов выполняют функции целостного организма. Деление клетки у одноклеточных влечет за собой увеличение количества особей, а в их жизненном цикле отсутствуют многоклеточные стадии. В целом у одноклеточных организмов совпадают клеточный и организменный уровни организации. Одноклеточными является подавляющее большинство бактерий, часть животных (простейшие), растений (некоторые водоросли) и грибов. Некоторые систематики даже предлагают выделить одноклеточные организмы в особое царство — протистов.
Колониальными называют организмы, у которых в процессе бесполого размножения дочерние особи остаются соединенными с материнским организмом, образуя более или менее сложное объединение — колонию. Кроме колоний многоклеточных организмов, таких как коралловые полипы, имеются и колонии одноклеточных, в частности водоросли пандорина и эвдорина. Колониальные организмы, по-видимому, были промежуточным звеном в процессе возникновения многоклеточных.
Многоклеточные организмы, вне всякого сомнения, обладают более высоким уровнем организации, чем одноклеточные, поскольку их тело образовано множеством клеток. В отличие от колониальных, которые также могут иметь более одной клетки, у многоклеточных организмов клетки специализируются на выполнении различных функций, что отражается и в их строении. Платой за эту специализацию является утрата их клетками способности к самостоятельному существованию, а зачастую и к воспроизведению себе подобных. Деление отдельной клетки приводит к росту многоклеточного организма, но не к его размножению. Онтогенез многоклеточных характеризуется процессом дробления оплодотворенной яйцеклетки на множество клеток-бластомеров, из которых в дальнейшем формируется организм с дифференцированными тканями и органами. Многоклеточные организмы, как правило, крупнее одноклеточных. Увеличение размеров тела по отношению к их поверхности способствовало усложнению и совершенствованию процессов обмена, формированию внутренней среды и, в конечном итоге, обеспечило им большую устойчивость к воздействиям окружающей среды (гомеостаз). Таким образом, многоклеточные обладают рядом преимуществ в организации по сравнению с одноклеточными и представляют собой качественный скачок в процессе эволюции. Многоклеточными являются немногие бактерии, большинство растений, животных и грибов.
Автотрофы и гетеротрофы
По способу питания все организмы делятся на автотрофов и гетеротрофов. Автотрофы способны самостоятельно синтезировать органические вещества из неорганических, а гетеротрофы используют исключительно готовые органические вещества.
Часть автотрофов может использовать для синтеза органических соединений энергию света — такие организмы называются фотоавтотрофами, они способны осуществлять фотосинтез. Фотоавтотрофами являются растения и часть бактерий. К ним тесно примыкают хемоавтотрофы, которые извлекают энергию путем окисления неорганических соединений в процессе хемосинтеза — это некоторые бактерии.
К гетеротрофам относятся как животные, так и грибы, бактерии и даже лишенные хлорофилла растения. Среди гетеротрофов имеются паразиты, сапротрофы, симбионты, хищники и т. д.
Паразиты — это организмы, использующие другие организмы (хозяев) в качестве среды обитания и источника питания. Характерными представителями этой группы гетеротрофов являются черви-паразиты кишечника человека — бычий цепень, острица и др.
Сапротрофами называют гетеротрофные организмы, осуществляющие питание органическими остатками. Они играют важную роль в круговороте веществ в природе, поскольку обеспечивают завершение существования органических веществ в природе, разлагая их до неорганических. Тем самым сапротрофы участвуют в процессах почвообразования, очистки вод и т. п. К сапротрофам относятся многие грибы и бактерии, а также некоторые растения и животные.
Симбионтами называют разноименные организмы, сосуществующие и взаимодействующие на различной основе. В широком смысле симбиозом называют не только взаимовыгодное сосуществование (мутуализм), как у человека с его бактериальной микрофлорой кишечника, но и негативное действие со стороны одного из партнеров — паразитизм.
Аэробы и анаэробы
По особенностям энергетического обмена организмы могут быть поделены на аэробов и анаэробов.
Аэробы способны жить и развиваться только при наличии в среде молекулярного кислорода, который они используют в качестве конечного акцептора электронов в процессе кислородного дыхания. К аэробам относится подавляющее большинство животных и грибов, все растения, а также значительная часть прокариот.
Анаэробы не используют кислород для осуществления процессов диссимиляции. Анаэробами являются некоторые животные (в основном внутренние паразиты), а также ряд бактерий. У животных-анаэробов функционирует главным образом гликолиз, а у бактерий — брожение, анаэробное (например, серное) дыхание и бескислородный фотосинтез. Наличие кислорода в среде не мешает развитию многих анаэробов.
Анаэробные организмы возникли раньше аэробных, так как в первичной атмосфере планеты не было кислорода. Его накопление связано с возникновением фотосинтеза, в связи с чем ряд организмов перешел к кислородному дыханию.
Воспроизведение организмов, его значение. Способы размножения, сходство и отличие полового и бесполого размножения. Оплодотворение у цветковых растений и позвоночных животных. Внешнее и внутреннее оплодотворение
Воспроизведение организмов, его значение
Способность организмов воспроизводить себе подобных является одним из фундаментальных свойств живого. Несмотря на то, что жизнь в целом непрерывна, продолжительность жизни отдельно взятой особи конечна, поэтому передача наследственной информации от одного поколения следующему при воспроизведении обеспечивает выживание данного вида организмов на протяжении длительных периодов времени. Таким образом, размножение обеспечивает непрерывность и преемственность жизни.
Обязательным условием воспроизведения является получение большего числа потомков, нежели родительских особей, поскольку далеко не все потомки смогут дожить до той стадии развития, на которой сами смогут давать потомство, так как они могут быть уничтожены хищниками, погибнуть от болезней и стихийных бедствий, например пожаров, наводнений и т. п.
Способы размножения, сходство и отличие полового и бесполого размножения
В природе различают два основных способа размножения — бесполое и половое.
Бесполое размножение — это способ размножения, при котором не происходит ни образования, ни слияния специализированных половых клеток — гамет, в нем принимает участие всего один родительский организм. В основе бесполого размножения лежит митотическое деление клетки.
В зависимости от того, сколько клеток материнского организма дает начало новой особи, бесполое размножение подразделяют на собственно бесполое и вегетативное. При собственно бесполом размножении дочерняя особь развивается из единственной клетки материнского организма, а при вегетативном — из группы клеток или целого органа.
В природе встречается четыре основных вида собственно бесполого размножения: бинарное деление, множественное деление, спорообразование и простое почкование.
Бинарное деление по сути представляет собой простое митотическое деление одноклеточного материнского организма, при котором вначале делится ядро, а затем и цитоплазма. Оно характерно для различных представителей растительного и животного царства, например амебы протей и инфузории-туфельки.
Множественному делению, или шизогонии, предшествует неоднократное деление ядра, после чего цитоплазма делится на соответствующее число фрагментов. Такой вид бесполого размножения встречается у одноклеточных животных — споровиков, например у малярийного плазмодия.
У многих растений и грибов в жизненном цикле происходит образование спор — одноклеточных специализированных образований, содержащих запас питательных веществ и покрытых плотной защитной оболочкой. Споры разносятся ветром и водой, и при наличии благоприятных условий прорастают, давая начало новому многоклеточному организму.
Характерным примером почкования как разновидности собственно бесполого размножения является почкование дрожжей, при котором на поверхности материнской клетки после деления ядра появляется небольшое выпячивание, в которое перемещается одно из ядер, после чего новая маленькая клетка отшнуровывается. Таким образом сохраняется способность материнской клетки к дальнейшему делению, а численность особей быстро увеличивается.
Вегетативное размножение может осуществляться в форме почкования, фрагментации, полиэмбрионии и др. При почковании у гидры образуется выпячивание стенки тела, которое постепенно увеличивается в размерах, на переднем конце прорывается ротовое отверстие, окруженное щупальцами. Завершается оно образованием маленькой гидры, которая отделяется затем от материнского организма. Почкование характерно также для ряда коралловых полипов и кольчатых червей.
Фрагментация сопровождается разделением тела на две и более части, причем из каждой развиваются полноценные особи (медузы, актинии, плоские и кольчатые черви, иглокожие).
При полиэмбрионии происходит разделение зародыша, сформировавшегося в том числе и в результате оплодотворения, на несколько зародышей. Такое явление регулярно происходит у броненосцев, но может происходить и у человека в случае однояйцевых близнецов.
Наиболее высоко развита способность к вегетативному размножению у растений, у которых начало новому организму могут давать клубни, луковицы, корневища, корневые отпрыски, усы и даже выводковые почки.
Для бесполого размножения требуется только одна родительская особь, что экономит время и энергию, необходимые на поиски полового партнера. Кроме того, из каждого фрагмента материнского организма могут возникнуть новые особи, что также является экономией вещества и энергии, затрачиваемых на размножение. Скорость бесполого размножения также достаточно велика, например, бактерии способны делиться каждые 20–30 минут, чрезвычайно быстро увеличивая свою численность. При этом способе размножения образуются генетически идентичные потомки — клоны, что может рассматриваться как преимущество при условии сохранения постоянства условий окружающей среды.
Однако в связи с тем, что единственным источником генетической изменчивости являются случайные мутации, практически полное отсутствие изменчивости среди потомков снижает их приспособляемость к новым условиям среды обитания при расселении и, как следствие, они погибают в гораздо больших количествах, нежели при половом размножении.
Половое размножение — способ размножения, при котором происходит образование и слияние половых клеток, или гамет, в одну клетку — зиготу, из которой развивается новый организм.
Если бы при половом размножении сливались соматические клетки с диплоидным набором хромосом (у человека 2n = 46), то уже во втором поколении в клетках нового организма содержался бы тетраплоидный набор (у человека 4n = 92), в третьем — октаплоидный и т. д.
Однако размеры эукариотической клетки не беспредельны, они должны колебаться в пределах 10–100 мкм, поскольку при меньших размерах клетки она не будет содержать полного набора необходимых для ее жизнедеятельности веществ и структур, а при больших размерах будет нарушаться равномерное обеспечение клетки кислородом, углекислым газом, водой и другими необходимыми веществами. Соответственно и размеры ядра, в котором находятся хромосомы, не могут превышать 1/5–1/10 объема клетки, а при нарушении этих условий клетка уже не сможет существовать. Таким образом, для полового размножения необходимо предварительное уменьшение количества хромосом, которое будет восстанавливаться при оплодотворении, что и обеспечивается процессом мейотического деления клетки.
Уменьшение числа хромосом должно быть к тому же строго упорядоченным и равноценным, поскольку, если новый организм не будет иметь полных пар хромосом при их общем нормальном количестве, то он либо не будет жизнеспособным, либо это будет сопровождаться развитием тяжелых заболеваний.
Таким образом, мейоз обеспечивает уменьшение числа хромосом, которое восстанавливается при оплодотворении, поддерживая в целом постоянство кариотипа.
Особыми формами полового размножения являются партеногенез и конъюгация. При партеногенезе, или девственном развитии, новый организм развивается из неоплодотворенной яйцеклетки, как, например, у дафний, медоносных пчел и некоторых скальных ящериц. Иногда этот процесс стимулируется внедрением сперматозоидов организмов другого вида.
В процессе конъюгации, которая характерна, например, для инфузорий, особи обмениваются фрагментами наследственной информации, а затем размножаются бесполым путем. Строго говоря, конъюгация является половым процессом, а не примером полового размножения.
Существование полового размножения требует выработки по меньшей мере двух видов половых клеток: мужских и женских. Животные организмы, у которых мужские и женские половые клетки вырабатываются разными особями, называются раздельнополыми, тогда как способные вырабатывать оба вида гамет — гермафродитами. Гермафродитизм характерен для многих плоских и кольчатых червей, брюхоногих моллюсков.
Растения, у которых мужские и женские цветки или другие разноименные половые органы располагаются на разных особях, называются двудомными, а имеющие одновременно оба вида цветков — однодомными.
Половое размножение обеспечивает возникновение генетического разнообразия потомков, основу которого составляют мейоз и рекомбинация родительских генов при оплодотворении. Наиболее удачные комбинации генов обеспечивают лучшее приспособление потомков к среде обитания, их выживание и большую вероятность передачи своей наследственной информации следующим поколениям. Этот процесс приводит к изменению признаков и свойств организмов и, в конечном итоге, к образованию новых видов в процессе эволюционного естественного отбора.
Вместе с тем вещество и энергия при половом размножении используются неэффективно, поскольку организмы вынуждены зачастую продуцировать миллионы гамет, однако только отдельные из них используются при оплодотворении. Кроме того, приходится затрачивать энергию и на обеспечение других условий. Например, растения образуют цветки и вырабатывают нектар, чтобы привлечь животных, которые переносят пыльцу на женские части других цветков, а животные затрачивают много времени и энергии на поиски брачных партнеров и ухаживание. Затем приходится расходовать много энергии на заботу о потомстве, поскольку при половом размножении потомки вначале зачастую бывают такими мелкими, что многие из них гибнут от хищников, голода или просто из-за неблагоприятных условий. Следовательно, при бесполом размножении затраты энергии гораздо меньше. Тем не менее половое размножение имеет по меньшей мере одно неоценимое достоинство — генетическую изменчивость потомства.
Бесполое и половое размножение широко используются человеком в сельском хозяйстве, декоративном животноводстве, растениеводстве и других областях для выведения новых сортов растений и пород животных, сохранения хозяйственно ценных признаков, а также быстрого увеличения числа особей.
При бесполом размножении растений, наряду с традиционными способами — черенкованием, прививкой и размножением отводками, постепенно занимают ведущее положение современные методы, связанные с использованием культуры тканей. При этом новые растения получают из небольших фрагментов материнского растения (клеток или кусочков ткани), выращенных на питательной среде, содержащей все необходимые растению питательные вещества и гормоны. Эти методы позволяют не только быстро размножить сорта растений с ценными признаками, например картофель, устойчивый к вирусу скручивания листьев, но и получить незараженные вирусами и другими возбудителями болезней растений организмы. Культура тканей лежит и в основе получения так называемых трансгенных, или генетически модифицированных организмов, а также гибридизации соматических клеток растений, которые невозможно скрестить иным путем.
Скрещивание растений различных сортов дает возможность получить организмы с новыми комбинациями хозяйственно ценных признаков. Для этого используют опыление пыльцой растений того же или другого вида и даже рода. Это явление называется отдаленной гибридизацией.
Поскольку у высших животных способность к естественному бесполому размножению отсутствует, основным способом их размножения является половое. Для этого применяют скрещивание особей как одного вида (породы), так и межвидовую гибридизацию, при этом получаются такие хорошо известные гибриды, как мул и лошак, в зависимости от того, особи какого вида были взяты в качестве материнских — осла и лошади. Однако межвидовые гибриды зачастую стерильны, то есть неспособны давать потомство, поэтому каждый раз их следует выводить заново.
Для размножения сельскохозяйственных животных используется и искусственный партеногенез. Выдающийся русский генетик Б. Л. Астауров, повышая температуру, вызвал больший выход самок тутового шелкопряда, которые плетут коконы из более тонкой и ценной нити, чем самцы.
Бесполым же размножением можно считать и клонирование, поскольку при этом используется ядро соматической клетки, которая вводится в оплодотворенную яйцеклетку с убитым ядром. Развивающийся организм должен быть копией, или клоном уже существующего организма.
Оплодотворение у цветковых растений и позвоночных животных
Оплодотворение — это процесс слияния мужских и женских половых клеток с образованием зиготы.
В процессе оплодотворения сначала происходит узнавание и физический контакт мужских и женских гамет, затем слияние их цитоплазмы, и только на последнем этапе объединение наследственного материала. Оплодотворение позволяет восстановить диплоидный набор хромосом, редуцированный в процессе формирования половых клеток.
Чаще всего в природе встречается оплодотворение мужскими половыми клетками другого организма, однако в целом ряде случаев возможно также и проникновение собственных сперматозоидов — самооплодотворение. С эволюционной точки зрения самооплодотворение является менее выгодным, так как при этом вероятность возникновения новых комбинаций генов минимальна. Поэтому даже у большинства гермафродитных организмов происходит перекрестное оплодотворение. Данный процесс присущ как растениям, так и животным, однако в его протекании у вышеупомянутых организмов имеется целый ряд отличий.
Так, у цветковых растений оплодотворению предшествует опыление — перенос пыльцы, содержащей мужские половые клетки — спермии — на рыльце пестика. Там она прорастает, образуя пыльцевую трубку с передвигающимися по ней двумя спермиями. Достигнув зародышевого мешка, один спермий сливается с яйцеклеткой с образованием зиготы, а другой — с центральной клеткой (2n), давая начало впоследствии запасающей ткани вторичного эндосперма. Такой способ оплодотворения получил название двойного оплодотворения.
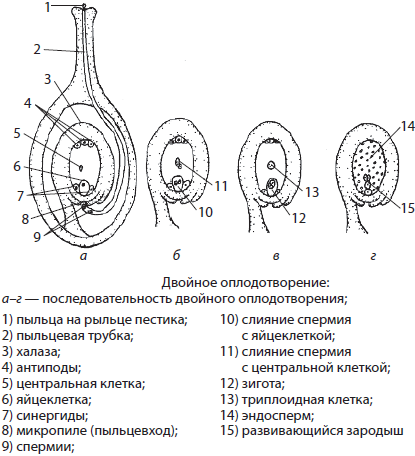
У животных, в частности позвоночных, оплодотворению предшествует сближение гамет, или осеменение. Успеху осеменения способствует синхронизация выведения мужских и женских половых клеток, а также выделение яйцеклетками специфических химических веществ с целью облегчения ориентации сперматозоидов в пространстве.
При разведении культурных растений и домашних животных усилия человека в основном направлены на сохранение и умножение хозяйственно ценных признаков, тогда как устойчивость этих организмов к условиям окружающей среды и жизнеспособность в целом снижаются. Кроме того, соя и многие другие культурные растения являются самоопыляемыми, поэтому для получения новых сортов необходимо вмешательство человека. Могут возникать также и затруднения в самом процессе оплодотворения, поскольку некоторые растения и животные могут иметь гены стерильности.
У растений для целей селекции производится искусственное опыление, для которого из цветков удаляют тычинки, а затем наносят на рыльца пестиков пыльцу из других цветков и накрывают опыленные цветки колпачками-изоляторами во избежание опыления пыльцой других растений. В некоторых случаях искусственное опыление производят для повышения урожайности, поскольку из завязей неопыленных цветков семена и плоды не развиваются. Такой прием практиковали ранее в посевах подсолнечника.
При отдаленной гибридизации, особенно если растения различаются по числу хромосом, естественное оплодотворение становится либо вовсе невозможным, либо уже при первом делении клетки происходит нарушение расхождения хромосом и организм гибнет. В таком случае оплодотворение производят в искусственных условиях, а в начале деления клетку обрабатывают колхицином — веществом, разрушающим веретено деления, при этом хромосомы рассыпаются по клетке, а затем формируется новое ядро уже с удвоенным числом хромосом, и при последующих делениях таких проблем не возникает. Таким образом были созданы редечно-капустный гибрид Г. Д. Карпеченко и тритикале — высокоурожайный гибрид пшеницы и ржи.
У основных видов сельскохозяйственных животных существует еще больше препятствий для оплодотворения, чем у растений, что вынуждает человека к применению решительных мер. Искусственное оплодотворение применяется в основном при разведении скота ценных пород, когда необходимо получить как можно больше потомства от одного производителя. В этих случаях семенную жидкость собирают, смешивают с водой, помещают в ампулы, а затем, по мере необходимости, вводят в половые пути самок. В рыбоводческих хозяйствах при искусственном оплодотворении у рыб полученную из молок сперму самцов смешивают с икрой в специальных емкостях. Молодь, выращенная в специальных садках, затем выпускается в естественные водоемы и восстанавливает популяцию, например, осетровых в Каспийском море и на Дону.
Таким образом, искусственное оплодотворение служит человеку для получения новых, высокопродуктивных сортов растений и пород животных, а также для повышения их продуктивности и восстановления природных популяций.
Внешнее и внутреннее оплодотворение
У животных различают внешнее и внутреннее оплодотворения. При внешнем оплодотворении женские и мужские половые клетки выводятся наружу, где и происходит процесс их слияния, как, например, у кольчатых червей, двустворчатых моллюсков, бесчерепных, большинства рыб и многих земноводных. Несмотря на то, что оно не требует сближения размножающихся особей, у подвижных животных возможно не только их сближение, но и скопление, как при нересте рыб.
Внутреннее оплодотворение связано с введением мужских половых продуктов в половые пути самки, и наружу выводится уже оплодотворенная яйцеклетка. Она зачастую имеет плотные оболочки, препятствующие ее повреждению и проникновению следующих сперматозоидов. Внутреннее оплодотворение характерно для подавляющего большинства наземных животных, например, для плоских и круглых червей, многих членистоногих и брюхоногих моллюсков, пресмыкающихся, птиц и млекопитающих, а также для ряда земноводных. Оно встречается и у некоторых водных животных, в том числе у головоногих моллюсков и хрящевых рыб.
Существует и промежуточный тип оплодотворения — наружно-внутренний, при котором самка захватывает половые продукты, специально оставленные самцом на каком-либо субстрате, как это происходит у некоторых членистоногих и хвостатых земноводных. Наружно-внутреннее оплодотворение может рассматриваться как переходное от внешнего к внутреннему.
Как внешнее, так и внутреннее оплодотворения имеют свои преимущества и недостатки. Так, при внешнем оплодотворении половые клетки выделяются в воду или воздух, вследствие чего подавляющее большинство их гибнет. Однако эта разновидность оплодотворения обеспечивает существование полового размножения у таких прикрепленных и малоподвижных животных, как двустворчатые моллюски и бесчерепные. При внутреннем оплодотворении потери гамет, безусловно, гораздо меньше, однако при этом вещество и энергия затрачиваются на поиск партнера, а появившиеся на свет потомки зачастую слишком малы и слабы и требуют длительной опеки родителей.
Онтогенез и присущие ему закономерности. Эмбриональное и постэмбриональное развитие организмов. Причины нарушения развития организмов
Онтогенез и присущие ему закономерности
Онтогенез (от греч. онтос — сущее и генезис — возникновение, происхождение) — это процесс индивидуального развития организма от зарождения до смерти. Данный термин был введен в 1866 году немецким ученым Э. Геккелем (1834–1919).
Зарождением организма считается возникновение зиготы в результате оплодотворения яйцеклетки сперматозоидом, хотя при партеногенезе зигота как таковая не образуется. В процессе онтогенеза происходят рост, дифференцировка и интеграция частей развивающегося организма. Дифференцировкой (от лат. дифферентио — различие) называют процесс возникновения различий между однородными тканями и органами, их изменения в ходе развития особи, приводящие к формированию специализированных тканей и органов.
Закономерности онтогенеза являются предметом изучения эмбриологии (от греч. эмбрион — зародыш и логос — слово, наука). Значительный вклад в ее развитие внесли русские ученые К. Бэр (1792–1876), обнаруживший яйцеклетку млекопитающих и положивший эмбриологические доказательства в основу классификации позвоночных животных, А. О. Ковалевский (1849–1901) и И. И. Мечников (1845–1916) — основоположники теории зародышевых листков и сравнительной эмбриологии, а также А. Н. Северцов (1866–1936), который выдвинул теорию возникновения новых признаков на любом этапе онтогенеза.
Индивидуальное развитие характерно только для многоклеточных организмов, поскольку у одноклеточных рост и развитие заканчиваются на уровне единственной клетки, а дифференцировка и вовсе отсутствует. Ход онтогенеза определяется генетическими программами, закрепившимися в процессе эволюции, то есть онтогенез является кратким повторением исторического развития данного вида, или филогенеза.
Несмотря на неизбежное переключение отдельных групп генов в ходе индивидуального развития, все изменения в организме происходят постепенно и не нарушают его целостности, однако события каждой предыдущей стадии оказывают значительное влияние на протекание последующих стадий развития. Так, любые сбои в процессе развития способны привести к прерыванию процесса онтогенеза на любой из стадий, как это достаточно часто происходит с зародышами (так называемые выкидыши).
Таким образом, для процесса онтогенеза характерно единство пространства и времени действия, поскольку он неразрывно связан с телом особи и протекает однонаправленно.
Эмбриональное и постэмбриональное развитие организмов
Периоды онтогенеза
Существует несколько периодизаций онтогенеза, однако чаще всего в онтогенезе животных выделяют эмбриональный и постэмбриональный периоды.
Эмбриональный период начинается с образования зиготы в процессе оплодотворения и заканчивается рождением организма или выходом его из зародышевых (яйцевых) оболочек.
Постэмбриональный период продолжается от рождения до смерти организма. Иногда выделяют и проэмбриональный период, или прогенез, к которому относят гаметогенез и оплодотворение. Эмбриональное развитие, или эмбриогенез, у животных и человека делят на ряд стадий: дробление, гаструляция, гистогенез и органогенез, а также период дифференцированного зародыша.
Дробление — это процесс митотического деления зиготы на все более мелкие клетки — бластомеры. Сначала образуются две клетки, затем четыре, восемь и т. д. Уменьшение размеров клеток связано в основном с тем, что в интерфазе клеточного цикла по разным причинам отсутствует G1-период, в котором должно происходить увеличение размеров дочерних клеток. Этот процесс похож на колку льда, однако является не хаотическим, а строго упорядоченным. Например, у человека это дробление является билатеральным, то есть двустороннесимметричным. В результате дробления и последующего расхождения клеток образуется бластула — однослойный многоклеточный зародыш, представляющий собой полый шарик, стенки которого образованы клетками — бластомерами, а полость внутри заполнена жидкостью и называется бластоцелем.
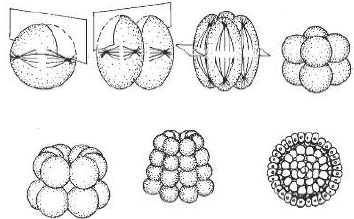
Гаструляцией называют процесс образования двух- или трехслойного зародыша — гаструлы (от греч. гастер — желудок), который происходит сразу после образования бластулы. Гаструляция осуществляется путем движения клеток и их групп относительно друг друга, например, впячиванием одной из стенок бластулы. Помимо двух или трех слоев клеток, гаструла имеет также первичный рот — бластопор.
Слои клеток гаструлы называются зародышевыми листками. Различают три зародышевых листка: эктодерму, мезодерму и энтодерму. Эктодерма (от греч. эктос — вне, снаружи и дерма — кожа) — это наружный зародышевый листок, мезодерма (от греч. мезос — средний, промежуточный) — средний, а энтодерма (от греч. энтос — внутри) — внутренний.
Несмотря на то, что все клетки развивающегося организма ведут свое происхождение от единственной клетки — зиготы — и содержат такой же набор генов, то есть являются ее клонами, поскольку образуются в результате митотического деления, процесс гаструляции сопровождается дифференцировкой клеток. Дифференцировка обусловлена переключением групп генов в различных частях зародыша и синтезом новых белков, определяющих в дальнейшем специфические функции клетки и накладывающих отпечаток на ее строение.
На специализацию клеток накладывает отпечаток и соседство других клеток, а также гормональный фон. Например, если от одного зародыша лягушки пересадить другому фрагмент, на котором развивается хорда, то это вызовет образование зачатка нервной системы в неположенном месте, и начнет формироваться как бы двойной зародыш. Это явление получило название эмбриональной индукции.
Гистогенезом называют процесс формирования зрелых тканей, присущих взрослому организму, а органогенезом — процесс формирования органов.
В процессе гисто- и органогенеза из эктодермы формируются эпидермис кожи и его производные (волосы, ногти, когти, перья), эпителий ротовой полости и эмаль зубов, прямая кишка, нервная система, органы чувств, жабры и др. Производными энтодермы являются кишечник и связанные с ним железы (печень и поджелудочная), а также легкие. А мезодерма дает начало всем видам соединительной ткани, в т. ч. костной и хрящевой тканям скелета, мышечной ткани скелетных мышц, кровеносной системе, многим эндокринным железам и т. д.
Закладка нервной трубки на спинной стороне зародыша хордовых животных символизирует начало еще одной промежуточной стадии развития — нейрулы (новолат. нейрула, уменьшит. от греч. нейрон — нерв). Этот процесс также сопровождается закладкой комплекса осевых органов, например хорды.
После протекания органогенеза наступает период дифференцированного зародыша, который характеризуется продолжением специализации клеток организма и быстрым ростом.
У многих животных в процессе эмбрионального развития возникают зародышевые оболочки и другие временные органы, которые не пригодятся в последующем развитии, например плацента, пуповина и др.
Постэмбриональное развитие животных по способности к репродукции делят на дорепродуктивный (ювенильный), репродуктивный и пострепродуктивный периоды.
Ювенильный период продолжается от рождения до полового созревания, он характеризуется интенсивным ростом и развитием организма.
По характеру развития различают прямое и непрямое развитие. При прямом развитии появляющийся на свет организм уже похож на взрослую особь, и процесс развития заключается в основном в увеличении линейных размеров особи, а также в формировании половых органов, как у человека. При непрямом развитии особь непохожа на взрослую, и в процессе развития происходит существенная перестройка ее организма, как у амфибий. Наличие в жизненном цикле личинок, непохожих на взрослых особей, позволяет снижать внутривидовую конкуренцию за счет разделения источников питания, способствует расселению малоподвижных или неподвижных организмов, обеспечивает заражение хозяев паразитами, а у некоторых организмов, которые не питаются на взрослой стадии, личинки выполняют и питательную функцию.
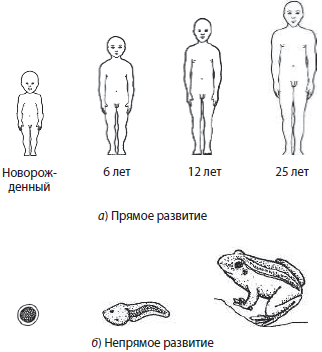
Рост организма происходит за счет увеличения количества клеток вследствие деления и увеличения их размеров. Выделяют два основных типа роста: ограниченный и неограниченный. Ограниченный, или закрытый рост происходит только в определенные периоды жизни, в основном до полового созревания. Он характерен для большинства животных. Например, человек растет в основном до 13–15 лет, хотя окончательное формирование тела происходит до 25 лет. Неограниченный, или открытый рост продолжается в течение всей жизни особи, как у растений и некоторых рыб. Также существуют периодический и непериодический рост.
Процессы роста контролирует эндокринная, или гормональная система: у человека увеличению линейных размеров тела способствует выделение соматотропного гормона, тогда как гонадотропные гормоны в значительной степени подав ляют его. Аналогичные механизмы открыты и у насекомых, у которых существует специальный ювенильный гормон и гормон линьки.
У цветковых растений эмбриональное развитие протекает после двойного оплодотворения, при котором один спермий оплодотворяет яйцеклетку, а второй — центральную клетку. Из зиготы образуется зародыш, который претерпевает ряд делений. После первого деления из одной клетки формируется собственно зародыш, а из второй — подвесок, через который происходит снабжение зародыша питательными веществами. Центральная клетка дает начало триплоидному эндосперму, содержащему питательные вещества для развития зародыша.
Эмбриональное и постэмбриональное развитие семенных растений зачастую разделены во времени, поскольку им требуются определенные условия для прорастания. Постэмбриональный период у растений делится на вегетативный, генеративный периоды и период старения. В вегетативном периоде происходит увеличение биомассы растения, в генеративном они приобретают способность к половому размножению (у семенных — к цветению и плодоношению), тогда как в период старения способность к репродукции утрачивается.
Жизненные циклы и чередование поколений
Вновь образовавшиеся организмы не сразу приобретают способность к воспроизведению себе подобных.
Жизненный цикл — совокупность стадий развития, начиная от зиготы, пройдя которые организм достигает зрелости и приобретает способность к размножению.
В жизненном цикле происходит чередование стадий развития с гаплоидным и диплоидным наборами хромосом, при этом у высших растений и животных преобладает диплоидный набор, а у низших — наоборот.
Жизненные циклы могут быть простыми и сложными. В отличие от простого жизненного цикла, в сложном половое размножение чередуется с партеногенетическим и бесполым. Например, рачки дафнии, дающие в течение лета бесполые поколения, осенью размножаются половым способом. Особенно сложны жизненные циклы некоторых грибов. У ряда животных чередование полового и бесполого поколений происходит регулярно, и такой жизненный цикл называется правильным. Он характерен, например, для ряда медуз.
Длительность жизненного цикла определяется числом поколений, развивающихся в течение года, или числом лет, на протяжении которых организм осуществляет свое развитие. Например, растения делят на однолетние и многолетние.
Знание жизненных циклов необходимо для генетического анализа, поскольку в гаплоидном и диплоидном состояниях различным образом выявляется действие генов: в первом случае имеются большие возможности для проявления всех генов, тогда как во втором некоторые гены не обнаруживаются.
Причины нарушения развития организмов
Способность к саморегуляции и к противостоянию вредным влияниям среды возникает у организмов не сразу. В течение эмбрионального и постэмбрионального развития, когда многие защитные системы организма еще не сформировались, организмы обычно уязвимы для действия повреждающих факторов. Поэтому у животных и растений зародыш защищен специальными оболочками или самим материнским организмом. Он либо снабжен специальной питающей тканью, либо получает питательные вещества непосредственно от материнского организма. Тем не менее изменение внешних условий может ускорить или затормозить развитие эмбриона и даже вызвать возникновение различных нарушений.
Факторы, вызывающие отклонения в развитии зародыша, называются тератогенными, или тератогенами. В зависимости от природы этих факторов их делят на физические, химические и биологические.
К физическим факторам относится, прежде всего, ионизирующая радиация, провоцирующая многочисленные мутации плода, которые могут быть несовместимыми с жизнью.
Химическими тератогенами являются тяжелые металлы, бензапирен, выбрасываемый автомобилями и промышленными предприятиями, фенолы, ряд лекарственных препаратов, алкоголь, наркотики и никотин.
Особо вредное влияние на развитие эмбриона человека оказывает употребление его родителями алкоголя, наркотиков, курение табака, поскольку алкоголь и никотин угнетают клеточное дыхание. Недостаточное снабжение зародыша кислородом приводит к тому, что в формирующихся органах образуется меньшее количество клеток, органы оказываются недоразвитыми. Особенно чувствительна к недостатку кислорода нервная ткань. Употребление будущей матерью алкоголя, наркотиков, курение табака, злоупотребление лекарствами часто приводит к необратимому повреждению эмбриона и последующему рождению детей с умственной отсталостью или врожденными уродствами.
Генетика, ее задачи. Наследственность и изменчивость — свойства организмов. Методы генетики. Основные генетические понятия и символика. Хромосомная теория наследственности. Современные представления о гене и геноме
Генетика, ее задачи
Успехи естествознания и клеточной биологии в XVIII–XIX веках позволили ряду ученых высказать предположения о существовании неких наследственных факторов, определяющих, например, развитие наследственных болезней, однако эти предположения не были подкреплены соответствующими доказательствами. Даже сформулированная Х. де Фризом в 1889 году теория внутриклеточного пангенеза, которая предполагала существование в ядре клетки неких «пангенов », определяющих наследственные задатки организма, и выход в протоплазму только тех из них, которые определяют тип клетки, не смогла изменить ситуацию, как и теория «зародышевой плазмы» А. Вейсмана, согласно которой приобретенные в процессе онтогенеза признаки не наследуются.
Лишь труды чешского исследователя Г. Менделя (1822–1884) стали основополагающим камнем современной генетики. Однако, несмотря на то, что его труды цитировались в научных изданиях, современники не обратили на них внимания. И лишь повторное открытие закономерностей независимого наследования сразу тремя учеными — Э. Чермаком, К. Корренсом и Х. де Фризом — вынудило научную общественность обратиться к истокам генетики.
Генетика — это наука, изучающая закономерности наследственности и изменчивости и методы управления ими.
Задачами генетики на современном этапе являются исследование качественных и количественных характеристик наследственного материала, анализ структуры и функционирования генотипа, расшифровка тонкой структуры гена и методов регуляции генной активности, поиск генов, вызывающих развитие наследственных болезней человека и методов их «исправления», создание нового поколения лекарственных препаратов по типу ДНК-вакцин, конструирование с помощью средств генной и клеточной инженерии организмов с новыми свойствами, которые могли бы производить необходимые человеку лекарственные препараты и продукты питания, а также полная расшифровка генома человека.
Наследственность и изменчивость — свойства организмов
Наследственность — это способность организмов передавать свои признаки и свойства в ряду поколений.
Изменчивость — свойство организмов приобретать новые признаки в течение жизни.
Признаки — это любые морфологические, физиологические, биохимические и иные особенности организмов, по которым одни из них отличаются от других, например цвет глаз. Свойствами же называют любые функциональные особенности организмов, в основе которых лежит определенный структурный признак или группа элементарных признаков.
Признаки организмов можно разделить на качественные и количественные. Качественные признаки имеют два-три контрастных проявления, которые называют альтернативными признаками, например голубой и карий цвет глаз, тогда как количественные (удойность коров, урожайность пшеницы) не имеют четко выраженных различий.
Материальным носителем наследственности является ДНК. У эукариот различают два типа наследственности: генотипическую и цитоплазматическую. Носители генотипической наследственности локализованы в ядре и далее речь пойдет именно о ней, а носителями цитоплазматической наследственности являются находящиеся в митохондриях и пластидах кольцевые молекулы ДНК. Цитоплазматическая наследственность передается в основном с яйцеклеткой, поэтому называется также материнской.
В митохондриях клеток человека локализовано небольшое количество генов, однако их изменение может оказывать существенное влияние на развитие организма, например приводить к развитию слепоты или постепенному снижению подвижности. Пластиды играют не менее важную роль в жизни растений. Так, в некоторых участках листа могут присутствовать бесхлорофильные клетки, что приводит, с одной стороны, к снижению продуктивности растения, а с другой — такие пестролистные организмы ценятся в декоративном озеленении. Воспроизводятся такие экземпляры в основном бесполым способом, так как при половом размножении чаще получаются обычные зеленые растения.
Методы генетики
1. Гибридологический метод, или метод скрещиваний, заключается в подборе родительских особей и анализе потомства. При этом о генотипе организма судят по фенотипическим проявлениям генов у потомков, полученных при определенной схеме скрещивания. Это старейший информативный метод генетики, который наиболее полно впервые применил Г. Мендель в сочетании со статистическим методом. Данный метод неприменим в генетике человека по этическим соображениям.
2. Цитогенетический метод основан на исследовании кариотипа: числа, формы и величины хромосом организма. Изучение этих особенностей позволяет выявить различные патологии развития.
3. Биохимический метод позволяет определять содержание различных веществ в организме, в особенности их избыток или недостаток, а также активность целого ряда ферментов.
4. Молекулярно-генетические методы направлены на выявление вариаций в структуре и расшифровку первичной последовательности нуклеотидов исследуемых участков ДНК. Они позволяют выявить гены наследственных болезней даже у эмбрионов, установить отцовство и т. д.
5. Популяционно-статистический метод позволяет определить генетический состав популяции, частоту определенных генов и генотипов, генетический груз, а также наметить перспективы развития популяции.
6. Метод гибридизации соматических клеток в культуре позволяет определить локализацию определенных генов в хромосомах при слиянии клеток различных организмов, например, мыши и хомяка, мыши и человека и т. д.
Основные генетические понятия и символика
Ген — это участок молекулы ДНК, или хромосомы, несущий информацию об определенном признаке или свойстве организма.
Некоторые гены могут оказывать влияние на проявление сразу нескольких признаков. Такое явление называется плейотропией. Например, ген, обусловливающий развитие наследственного заболевания арахнодактилии (паучьи пальцы), вызывает также искривление хрусталика, патологии многих внутренних органов.
Каждый ген занимает в хромосоме строго определенное место — локус. Так как в соматических клетках большинства эукариотических организмов хромосомы парные (гомологичные), то в каждой из парных хромосом находится по одной копии гена, отвечающего за определенный признак. Такие гены называются аллельными.
Аллельные гены чаще всего существуют в двух вариантах — доминантном и рецессивном. Доминантной называют аллель, которая проявляется вне зависимости от того, какой ген находится в другой хромосоме, и подавляет развитие признака, кодируемого рецессивным геном. Доминантные аллели обозначаются обычно прописными буквами латинского алфавита (A, B, C и др.), а рецессивные — строчными (a, b, c и др.). Рецессивные аллели могут проявляться только в том случае, если они занимают локусы в обеих парных хромосомах.
Организм, у которого в обеих гомологичных хромосомах находятся одинаковые аллели, называется гомозиготным по данному гену, или гомозиготой (AA, aa, ААBB, ааbb и т. д.), а организм, у которого в обеих гомологичных хромосомах находятся разные варианты гена — доминантный и рецессивный — называется гетерозиготным по данному гену, или гетерозиготой (Aa, АаBb и т. д.).
Ряд генов может иметь три и более структурных варианта, например группы крови по системе AB0 кодируются тремя аллелями — IA, IB, i. Такое явление называется множественным аллелизмом. Однако даже в этом случае каждая хромосома из пары несет только одну аллель, то есть все три варианта гена у одного организма не могут быть представлены.
Геном — совокупность генов, характерная для гаплоидного набора хромосом.
Генотип — совокупность генов, характерная для диплоидного набора хромосом.
Фенотип — совокупность признаков и свойств организма, которая является результатом взаимодействия генотипа и окружающей среды.
Поскольку организмы отличаются между собой многими признаками, установить закономерности их наследования можно только при анализе двух и более признаков в потомстве. Скрещивание, при котором рассматривается наследование и проводится точный количественный учет потомства по одной паре альтернативных признаков, называется моногибридным, по двум парам — дигибридным, по большему количеству признаков — полигибридным.
По фенотипу особи далеко не всегда можно установить ее генотип, поскольку как гомозиготный по доминантному гену организм (АА), так и гетерозиготный (Аа) будет иметь в фенотипе проявление доминантной аллели. Поэтому для проверки генотипа организма с перекрестным оплодотворением применяют анализирующее скрещивание — скрещивание, при котором организм с доминантным признаком скрещивается с гомозиготным по рецессивному гену. При этом гомозиготный по доминантному гену организм не будет давать расщепления в потомстве, тогда как в потомстве гетерозиготных особей наблюдается равное количество особей с доминантным и рецессивным признаками.
Для записи схем скрещиваний чаще всего применяются следующие условные обозначения:
Р (от лат. парента — родители) — родительские организмы;
$♀$ (алхимический знак Венеры — зеркало с ручкой) — материнская особь;
$♂$ (алхимический знак Марса — щит и копье) — отцовская особь;
$×$ — знак скрещивания;
F1, F2, F3 и т. д. — гибриды первого, второго, третьего и последующих поколений;
Fа — потомство от анализирующего скрещивания.
Хромосомная теория наследственности
Основоположник генетики Г. Мендель, равно как и его ближайшие последователи, не имели ни малейшего представления о материальной основе наследственных задатков, или генов. Однако уже в 1902–1903 годах немецкий биолог Т. Бовери и американский студент У. Сэттон независимо друг от друга предположили, что поведение хромосом при созревании клеток и оплодотворении позволяет объяснить расщепление наследственных факторов по Менделю, т. е., по их мнению, гены должны быть расположены в хромосомах. Данные предположения стали краеугольным камнем хромосомной теории наследственности.
В 1906 году английские генетики У. Бэтсон и Р. Пеннет обнаружили нарушение менделевского расщепления при скрещивании душистого горошка, а их соотечественник Л. Донкастер в экспериментах с бабочкой крыжовенной пяденицей открыл сцепленное с полом наследование. Результаты этих экспериментов явно противоречили менделевским, но если учесть, что к тому времени уже было известно о том, что количество известных признаков для экспериментальных объектов намного превышало количество хромосом, а это наводило на мысль, что каждая хромосома несет более одного гена, а гены одной хромосомы наследуются совместно.
В 1910 году начинаются эксперименты группы Т. Моргана на новом экспериментальном объекте — плодовой мушке дрозофиле. Результаты этих экспериментов позволили к середине 20-х годов XX века сформулировать основные положения хромосомной теории наследственности, определить порядок расположения генов в хромосомах и расстояния между ними, т. е. составить первые карты хромосом.
Основные положения хромосомной теории наследственности:
- Гены расположены в хромосомах. Гены одной хромосомы наследуются совместно, или сцепленно, и называются группой сцепления. Число групп сцепления численно равно гаплоидному набору хромосом.
- Каждый ген занимает в хромосоме строго определенное место — локус.
- Гены в хромосомах расположены линейно.
- Нарушение сцепления генов происходит только в результате кроссинговера.
- Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними.
- Независимое наследование характерно только для генов негомологичных хромосом.
Современные представления о гене и геноме
В начале 40-х годов ХХ века Дж. Бидл и Э. Тейтум, анализируя результаты генетических исследований, проведенных на грибе нейроспоре, пришли к выводу, что каждый ген контролирует синтез какого-либо фермента, и сформулировали принцип «один ген — один фермент».
Однако уже в 1961 году Ф. Жакобу, Ж. Л. Моно и А. Львову удалось расшифровать структуру гена кишечной палочки и исследовать регуляцию его активности. За это открытие им в 1965 году была присуждена Нобелевская премия по физиологии и медицине.
В процессе исследования, кроме структурных генов, контролирующих развитие определенных признаков, им удалось выявить и регуляторные, основной функцией которых является проявление признаков, кодируемых другими генами.
Структура прокариотического гена. Структурный ген прокариот имеет сложное строение, поскольку в его состав входят регуляторные участки и кодирующие последовательности. К регуляторным участкам относятся промотор, оператор и терминатор. Промотором называют участок гена, к которому прикрепляется фермент РНК-полимераза, обеспечивающий синтез иРНК в процессе транскрипции. С оператором, располагающимся между промотором и структурной последовательностью, может связываться белок-репрессор, не позволяющий РНК-полимеразе начать считывание наследственной информации с кодирующей последовательности, и только его удаление позволяет начать транскрипцию. Структура репрессора закодирована обычно в регуляторном гене, находящемся в другом участке хромосомы. Считывание информации заканчивается на участке гена, который называется терминатором.
Кодирующая последовательность структурного гена содержит информацию о последовательности аминокислот в соответствующем белке. Кодирующую последовательность у прокариот называют цистроном, а совокупность кодирующих и регуляторных участков гена прокариот — опероном. В целом прокариоты, к которым относится и кишечная палочка, имеют сравнительно небольшое количество генов, расположенных в единственной кольцевой хромосоме.

Цитоплазма прокариот может содержать также дополнительные небольшие кольцевые или незамкнутые молекулы ДНК, которые называются плазмидами. Плазмиды способны встраиваться в хромосомы и передаваться от одной клетки к другой. Они могут нести информацию о половых признаках, патогенности и устойчивости к антибиотикам.
Структура эукариотического гена. В отличие от прокариот, гены эукариот не имеют оперонной структуры, поскольку не содержат оператора, и каждый структурный ген сопровождается только промотором и терминатором. Кроме того, в генах эукариот значащие участки (экзоны) чередуются с незначащими (интронами), которые полностью переписываются на иРНК, а затем вырезаются в процессе их созревания. Биологическая роль интронов состоит в снижении вероятности мутаций в значащих участках. Регуляция генов эукариот намного сложнее, нежели описанная для прокариот.
Геном человека. В каждой клетке человека в 46 хромосомах находится около 2 м ДНК, плотно упакованной в двойную спираль, которая состоит примерно из 3,2 $×$ 109 нуклеотидных пар, что обеспечивает около 101900000000 возможных уникальных комбинаций. К концу 80-х годов ХХ века было известно расположение примерно 1500 генов человека, однако их общее количество оценивали примерно в 100 тыс., поскольку только наследственных болезней у человека имеется примерно 10 тыс., не говоря уже о количестве разнообразных белков, содержащихся в клетках.
В 1988 году стартовал международный проект «Геном человека», который к началу XXI века закончился полной расшифровкой последовательности нуклеотидов. Он дал возможность понять, что два разных человека на 99,9 % имеют сходные последовательности нуклеотидов, и лишь остающиеся 0,1 % определяют нашу индивидуальность. Всего было обнаружено примерно 30–40 тыс. структурных генов, однако затем их количество было снижено до 25–30 тыс. Среди этих генов имеются не только уникальные, но и повторяющиеся сотни и тысячи раз. Тем не менее данные гены кодируют гораздо большее количество белков, например десятки тысяч защитных белков — иммуноглобулинов.
97 % нашего генома является генетическим «мусором», который существует только потому, что умеет хорошо воспроизводиться (РНК, которые транскрибируются на этих участках, никогда не покидают ядро). Например, среди наших генов есть не только «человеческие» гены, но и 60 % генов, похожих на гены мушки дрозофилы, а с шимпанзе нас роднит до 99 % генов.
Параллельно с расшифровкой генома происходило и картирование хромосом, вследствие этого удалось не только обнаружить, но и определить расположение некоторых генов, отвечающих за развитие наследственных заболеваний, а также генов-мишеней лекарственных препаратов.
Расшифровка генома человека пока не дает прямого эффекта, поскольку мы получили своеобразную инструкцию по сборке такого сложного организма, как человек, но не научились изготавливать его или хотя бы исправлять погрешности в нем. Тем не менее эра молекулярной медицины уже на пороге, во всем мире идет разработка так называемых генопрепаратов, которые смогут блокировать, удалять или даже замещать патологические гены у живых людей, а не только в оплодотворенной яйцеклетке.
Не следует забывать и о том, что в эукариотических клетках ДНК содержится не только в ядре, но также в митохондриях и пластидах. В отличие от ядерного генома, организация генов митохондрий и пластид имеет много общего с организацией генома прокариот. Несмотря на то что эти органеллы несут менее 1 % наследственной информации клетки и не кодируют даже полного набора белков, необходимых для их собственного функционирования, они способны существенно влиять на некоторые признаки организма. Так, пестролистность у растений хлорофитума, плюща и других наследует незначительное число потомков даже при скрещивании двух пестролистных растений. Это обусловлено тем, что пластиды и митохондрии передаются большей частью с цитоплазмой яйцеклетки, поэтому такая наследственность называется материнской, или цитоплазматической, в отличие от генотипической, которая локализуется в ядре.
Закономерности наследственности, их цитологические основы. Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание). Законы Т. Моргана: сцепленное наследование признаков, нарушение сцепления генов. Генетика пола. Наследование признаков, сцепленных с полом. Взаимодействие генов. Генотип как целостная система. Генетика человека. Методы изучения генетики человека. Решение генетических задач. Составление схем скрещивания
Закономерности наследственности, их цитологические основы
Согласно хромосомной теории наследственности каждая пара генов локализована в паре гомологичных хромосом, причем каждая из хромосом несет только по одному из этих факторов. Если представить, что гены являются точечными объектами на прямых — хромосомах, то схематически гомозиготные особи могут быть записаны как A||A или a||a, тогда как гетерозиготная — A||a. При образовании гамет в процессе мейоза каждый из генов пары гетерозиготы окажется в одной из половых клеток.
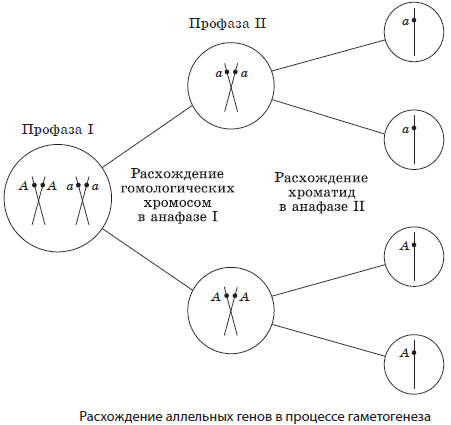
Например, если скрестить двух гетерозиготных особей, то при условии образования у каждой из них только пары гамет возможно получение всего лишь четырех дочерних организмов, три из которых будут нести хотя бы один доминантный ген А, и только один будет гомозиготен по рецессивному гену а, т. е. закономерности наследственности носят статистический характер.
В тех случаях, если гены располагаются в разных хромосомах, то при образовании гамет распределение между ними аллелей из данной пары гомологичных хромосом происходит совершенно независимо от распределения аллелей из других пар. Именно случайное расположение гомологичных хромосом на экваторе веретена в метафазе I мейоза и их последующее расхождение в анафазе I ведет к разнообразию рекомбинаций аллелей в гаметах.
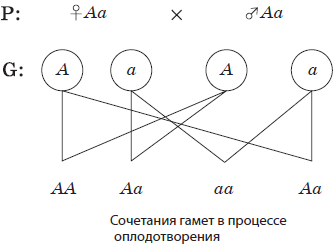
Число возможных сочетаний аллелей в мужских или женских гаметах можно определить по общей формуле 2n, где n — число хромосом, характерное для гаплоидного набора. У человека n = 23, а возможное число сочетаний составляет 223 = 8388608. Последующее объединение гамет при оплодотворении является также случайным, и поэтому в потомстве можно зафиксировать независимое расщепление по каждой паре признаков.
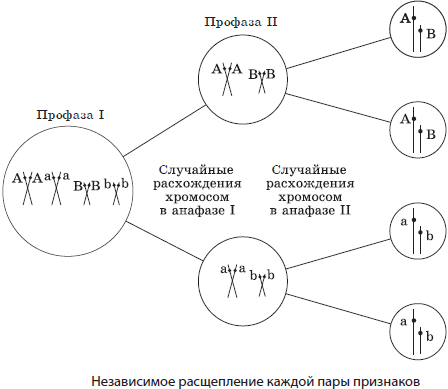
Однако число признаков у каждого организма во много раз больше числа его хромосом, которые можно различить под микроскопом, следовательно, каждая хромосома должна содержать множество факторов. Если представить себе, что у некоторой особи, гетерозиготной по двум парам генов, расположенных в гомологичных хромосомах, образуются гаметы, то следует учитывать не только вероятность образования гамет с исходными хромосомами, но и гамет, получивших измененные в результате кроссинговера в профазе I мейоза хромосомы. Следовательно, в потомстве возникнут новые сочетания признаков. Данные, полученные в экспериментах на дрозофиле, легли в основу хромосомной теории наследственности.
Другое фундаментальное подтверждение цитологической основы наследственности было получено при исследовании различных заболеваний. Так, у человека одна из форм рака обусловлена утратой маленького участка одной из хромосом.
Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание)
Основные закономерности независимого наследования признаков были открыты Г. Менделем, который достиг успеха, применив в своих исследованиях новый на тот момент гибридологический метод.
Успех Г. Менделя был обеспечен следующими факторами:
- удачным выбором объекта исследования (гороха посевного), который имеет короткий срок вегетации, является самоопыляемым растением, дает значительное количество семян и представлен большим количеством сортов с хорошо различимыми признаками;
- использованием только чистых линий гороха, которые на протяжении нескольких поколений не давали расщепления признаков в потомстве;
- концентрацией только на одном-двух признаках;
- планированием эксперимента и составлением четких схем скрещивания;
- точным количественным подсчетом полученного потомства.
Для исследования Г. Мендель отобрал только семь признаков, имеющих альтернативные (контрастные) проявления. Уже в первых скрещиваниях он обратил внимание, что в потомстве первого поколения при скрещивании растений с желтыми и зелеными семенами все потомство имело желтые семена. Аналогичные результаты были получены и при исследовании других признаков. Признаки, которые преобладали в первом поколении, Г. Мендель назвал доминантными. Те же из них, которые не проявлялись в первом поколении, получили название рецессивных.
Особи, которые давали расщепление в потомстве, получили название гетерозиготных, а особи, не дававшие расщепления, — гомозиготных.
Признаки гороха, наследование которых изучено Г. Менделем
| Признак | Вариант проявления | |
| Доминантный | Рецессивный | |
| Окраска семян | Желтая | Зеленая |
| Форма семян | Гладкая | Морщинистая |
| Форма плода (боба) | Простой | Членистый |
| Окраска плода | Зеленая | Желтая |
| Окраска венчика цветка | Красная | Белая |
| Положение цветков | Пазушные | Верхушечные |
| Длина стебля | Длинный | Короткий |
Скрещивание, при котором исследуется проявление только одного признака, называется моногибридным. В таком случае прослеживаются закономерности наследования только двух вариантов одного признака, развитие которых обусловлено парой аллельных генов. Например, признак «окраска венчика цветка» у гороха имеет только два проявления — красная и белая. Все остальные признаки, свойственные данным организмам, во внимание не принимаются и не учитываются в расчетах.
Схема моногибридного скрещивания такова:

Скрестив два растения гороха, одно из которых имело желтые семена, а другое — зеленые, в первом поколении Г. Мендель получал растения исключительно с желтыми семенами, независимо от того, какое растение было выбрано в качестве материнского, а какое — отцовского. Такие же результаты были получены и в скрещиваниях по другим признакам, что дало Г. Менделю основания сформулировать закон единообразия гибридов первого поколения, который также называют первым законом Менделя и законом доминирования.
Первый закон Менделя:
При скрещивании гомозиготных родительских форм, отличающихся по одной паре альтернативных признаков, все гибриды первого поколения будут единообразны как по генотипу, так и по фенотипу.
А — желтые семена; а — зеленые семена.

При самоопылении (скрещивании) гибридов первого поколения оказалось, что 6022 семени имеют желтую окраску, а 2001 — зеленую, что примерно соответствует соотношению 3:1. Обнаруженная закономерность получила название закона расщепления, или второго закона Менделя.
Второй закон Менделя:
При скрещивании гетерозиготных гибридов первого поколения в потомстве будет наблюдаться преобладание одного из признаков в соотношении 3:1 по фенотипу (1:2:1 по генотипу).

Однако по фенотипу особи далеко не всегда удается установить ее генотип, поскольку как гомозиготы по доминантному гену (АА), так и гетерозиготы (Аа) будут иметь в фенотипе проявление доминантного гена. Поэтому для организмов с перекрестным оплодотворением применяют анализирующее скрещивание — скрещивание, при котором организм с неизвестным генотипом скрещивается с гомозиготой по рецессивному гену для проверки генотипа. При этом гомозиготные особи по доминантному гену расщепления в потомстве не дают, тогда как в потомстве гетерозиготных наблюдается равное количество особей как с доминантным, так и с рецессивным признаками:

Основываясь на результатах собственных экспериментов, Г. Мендель предположил, что наследственные факторы при образовании гибридов не смешиваются, а сохраняются в неизменном виде. Поскольку связь между поколениями осуществляется через гаметы, то он допустил, что в процессе их образования в каждую из гамет попадает только один фактор из пары (т. е. гаметы генетически чисты), а при оплодотворении пара восстанавливается. Эти предположения получили название правила чистоты гамет.
Правило чистоты гамет:
При гаметогенезе гены одной пары разделяются, т. е. каждая гамета несет только один вариант гена.
Однако организмы отличаются друг от друга по многим признакам, поэтому установить закономерности их наследования возможно только при анализе двух и более признаков в потомстве.
Скрещивание, при котором рассматривается наследование и производится точный количественный учет потомства по двум парам признаков, называется дигибридным. Если же анализируется проявление большего числа наследственных признаков, то это уже полигибридное скрещивание.
Схема дигибридного скрещивания:
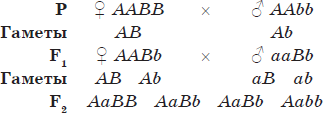
При большем разнообразии гамет определение генотипов потомков становится затруднительным, поэтому для анализа широко используется решетка Пеннета, в которую по горизонтали заносятся мужские гаметы, а по вертикали — женские. Генотипы потомков определяются сочетанием генов в столбцах и строках.
| $♀$/$♂$ | aB | ab |
| AB | AaBB | AaBb |
| Ab | AaBb | Aabb |
Для дигибридного скрещивания Г. Мендель выбрал два признака: окраску семян (желтую и зеленую) и их форму (гладкую и морщинистую). В первом поколении соблюдался закон единообразия гибридов первого поколения, а во втором поколении было 315 желтых гладких семян, 108 — зеленых гладких, 101 — желтое морщинистое и 32 зеленых морщинистых. Подсчет показал, что расщепление приближалось к 9:3:3:1, но по каждому из признаков сохранялось соотношение 3:1 (желтые — зеленые, гладкие — морщинистые). Эта закономерность получила название закона независимого расщепления признаков, или третьего закона Менделя.
Третий закон Менделя:
При скрещивании гомозиготных родительских форм, отличающихся по двум и более парам признаков, во втором поколении будет происходить независимое расщепление данных признаков в соотношении 3:1 (9:3:3:1 при дигибридном скрещивании).

| $♀$/$♂$ | AB | Ab | aB | ab |
| AB | AABB | AABb | AaBB | AaBb |
| Ab | AABb | AAbb | AaBb | Aabb |
| aB | AaBB | AaBb | aaBB | aaBb |
| ab | AaBb | Aabb | aaBb | aabb |
$F_2 {9A_B_}↙{\text"желтые гладкие"} : {3A_bb}↙{\text"желтые морщинистые"} : {3aaB_}↙{\text"зеленые гладкие"} : {1aabb}↙{\text"зеленые морщинистые"}$
Третий закон Менделя применим только к случаям независимого наследования, когда гены расположены в разных парах гомологичных хромосом. В тех случаях, когда гены расположены в одной паре гомологичных хромосом, действительны закономерности сцепленного наследования. Закономерности независимого наследования признаков, установленные Г. Менделем, также часто нарушаются и при взаимодействии генов.
Законы Т. Моргана: сцепленное наследование признаков, нарушение сцепления генов
Новый организм получает от родителей не россыпь генов, а целые хромосомы, при этом количество признаков и соответственно определяющих их генов гораздо больше, чем хромосом. В соответствии с хромосомной теорией наследственности, гены, расположенные в одной хромосоме, наследуются сцепленно. Вследствие этого при дигибридном скрещивании они не дают ожидаемого расщепления 9:3:3:1 и не подчиняются третьему закону Менделя. Можно было бы ожидать, что сцепление генов является полным, и при скрещивании гомозиготных по данным генам особей и во втором поколении дает исходные фенотипы в соотношении 3:1, а при анализирующем скрещивании гибридов первого поколения расщепление должно составлять 1:1.
Для проверки этого предположения американский генетик Т. Морган выбрал у дрозофилы пару генов, контролирующих окраску тела (серое — черное) и форму крыла (длинные — зачаточные), которые расположены в одной паре гомологичных хромосом. Серое тело и длинные крылья являются доминантными признаками. При скрещивании гомозиготной мухи с серым телом и длинными крыльями и гомозиготной мухи с черным телом и зачаточными крыль ями во втором поколении действительно были получены в основном родительские фенотипы в соотношении, близком к 3:1, однако имелось и незначительное количество особей с новыми комбинациями этих признаков. Данные особи называются рекомбинантными.
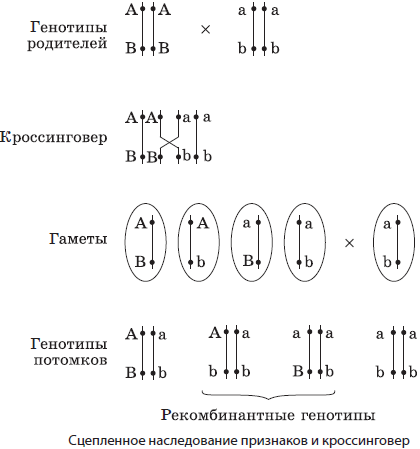
Однако, проведя анализирующее скрещивание гибридов первого поколения с гомозиготами по рецессивным генам, Т. Морган обнаружил, что 41,5 % особей имели серое тело и длинные крылья, 41,5 % — черное тело и зачаточные крылья, 8,5 % — серое тело и зачаточные крылья, и 8,5 % — черное тело и зачаточные крылья. Он связал полученное расщепление с кроссинговером, происходящим в профазе I мейоза и предложил считать единицей расстояния между генами в хромосоме 1 % кроссинговера, впоследствии названный в его честь морганидой.
Закономерности сцепленного наследования, установленные в ходе экспериментов на дрозофиле, получили название закона Т. Моргана.
Закон Моргана:
Гены, локализованные в одной хромосоме, занимают определенное место, называемое локусом, и наследуются сцепленно, причем сила сцепления обратно пропорциональна расстоянию между генами.
Гены, расположенные в хромосоме непосредственно друг за другом (вероятность кроссинговера крайне мала), называются сцепленными полностью, а если между ними находится еще хотя бы один ген, то они сцеплены не полностью и их сцепление нарушается при кроссинговере в результате обмена участками гомологичных хромосом.
Явления сцепления генов и кроссинговера позволяют построить карты хромосом с нанесенным на них порядком расположения генов. Генетические карты хромосом созданы для многих генетически хорошо изученных объектов: дрозофилы, мыши, человека, кукурузы, пшеницы, гороха и др. Изучение генетических карт позволяет сравнивать строение генома у различных видов организмов, что имеет важное значение для генетики и селекции, а также эволюционных исследований.
Генетика пола
Пол — это совокупность морфологических и физиологических особенностей организма, обеспечивающих половое размножение, сущность которого сводится к оплодотворению, то есть слиянию мужских и женских половых клеток в зиготу, из которой развивается новый организм.
Признаки, по которым один пол отличается от другого, делят на первичные и вторичные. К первичным половым признакам относятся половые органы, а все остальные — ко вторичным.
У человека вторичными половыми признаками являются тип телосложения, тембр голоса, преобладание мышечной или жировой ткани, наличие оволосения на лице, кадыка, молочных желез. Так, у женщин таз обычно шире плеч, преобладает жировая ткань, выражены молочные железы, голос — высокий. Мужчины же отличаются от них более широкими плечами, преобладанием мышечной ткани, наличием оволосения на лице и кадыка, а также низким голосом. Человечество издавна интересовал вопрос, почему особи мужского и женского пола рождаются в соотношении приблизительно 1:1. Объяснение этому было получено при изучении кариотипов насекомых. Оказалось, что у самок некоторых клопов, кузнечиков и бабочек на одну хромосому больше, чем у самцов. В свою очередь самцы продуцируют гаметы, различающиеся по количеству хромосом, тем самым заранее определяя пол потомка. Однако впоследствии было выяснено, что у большинства организмов количество хромосом у самцов и самок все же не различается, но у одного из полов есть пара хромосом, которые не подходят друг другу по размерам, а у другого все хромосомы парные.
В кариотипе человека также обнаружили подобное различие: у мужчин есть две непарные хромосомы. По форме эти хромосомы в начале деления напоминают латинские буквы Х и Y, и поэтому были названы Х- и Y-хромосомами. Сперматозоиды мужчины могут нести одну из этих хромосом и определять пол будущего ребенка. В связи с этим хромосомы человека и многих других организмов делят на две группы: аутосомы и гетерохромосомы, или половые хромосомы.
К аутосомам относят хромосомы, одинаковые для обоих полов, тогда как половые хромосомы — это хромосомы, отличающиеся у разных полов и несущие информацию о половых признаках. В тех случаях, когда пол несет одинаковые половые хромосомы, например ХХ, говорят, что он гомозиготен, или гомогаметен (образует одинаковые гаметы). Другой же пол, имеющий разные половые хромосомы (XY), называется гемизиготным (не имеющим полного эквивалента аллельных генов), или гетерогаметным. У человека, большинства млекопитающих, мушки дрозофилы и других организмов гомогаметен женский пол (ХХ), а мужской — гетерогаметен (XY), тогда как у птиц гомогаметен мужской пол (ZZ, или XX), а женский — гетерогаметен (ZW, или XY).
Х-хромосома является крупной неравноплечей хромосомой, которая несет свыше 1500 генов, причем многие мутантные их аллели вызывают у человека развитие тяжелых наследственных заболеваний, таких как гемофилия и дальтонизм. Y-хромосома, напротив, очень маленькая, в ней содержится всего около десятка генов, в том числе специфические гены, ответственные за развитие по мужскому типу.
Кариотип мужчины записывается как $♂$ 46, XY, а кариотип женщины — как $♀$ 46, ХХ.
Поскольку гаметы с половыми хромосомами продуцируются у самцов с равной вероятностью, то ожидаемое соотношение полов в потомстве составляет 1:1, что и совпадает с фактически наблюдаемым.
Пчелы отличаются от других организмов тем, что самки у них развиваются из оплодотворенных яиц, а самцы — из неоплодотворенных. Соотношение полов у них отличается от указанного выше, так как процесс оплодотворения регулирует матка, в половых путях которой с весны запасаются сперматозоиды на весь год.
У ряда организмов пол может определяться иным способом: до оплодотворения или после него, в зависимости от условий внешней среды.
Наследование признаков, сцепленных с полом
Так как некоторые гены находятся в половых хромосомах, неодинаковых у представителей противоположных полов, то характер наследования признаков, кодируемых данными генами, отличается от общего. Такой тип наследования называется крис-кросс наследованием, поскольку мужчины наследуют признаки матери, а женщины — отца. Признаки, определяемые генами, которые находятся в половых хромосомах, называются сцепленными с полом. Примерами признаков, сцепленных с полом, являются рецессивные признаки гемофилии и дальтонизма, которые в основном проявляются у мужчин, так как в Y-хромосоме нет аллельных генов. Женщины болеют такими болезнями только в том случае, если и от отца, и от матери они получили такие признаки.
Например, если мать была гетерозиготным носителем гемофилии, то у половины ее сыновей свертываемость крови будет нарушена:
ХH — нормальное свертывание крови
Хh — несвертываемость крови (гемофилия)

Признаки, закодированные в генах Y-хромосомы, передаются сугубо по мужской линии и называются голандрическими (наличие перепонки между пальцами ног, повышенное оволосение края ушной раковины).
Взаимодействие генов
Проверка закономерностей независимого наследования на различных объектах уже в начале XX века показала, что, например, у ночной красавицы при скрещивании растений с красным и белым венчиком у гибридов первого поколения венчики окрашены в розовый цвет, тогда как во втором поколении имеются особи с красными, розовыми и белыми цветками в соотношении 1:2:1. Это навело исследователей на мысль, что аллельные гены могут оказывать определенное влияние друг на друга. Впоследствии было также установлено, что и неаллельные гены способствуют проявлению признаков других генов или подавляют их. Данные наблюдения стали основой представления о генотипе как о системе взаимодействующих генов. В настоящее время различают взаимодействие аллельных и неаллельных генов.
Ко взаимодействию аллельных генов относят полное и неполное доминирование, кодоминирование и сверхдоминирование. Полным доминированием считают все случаи взаимодействия аллельных генов, при которых в гетерозиготе наблюдается проявление исключительно доминантного признака, как, например, окраска и форма семени у гороха.
Неполное доминирование — это тип взаимодействия аллельных генов, при котором проявление рецессивного аллеля в большей или меньшей степени ослабляет проявление доминантного, как в случае окраски венчика ночной красавицы (белая + красная = розовая) и шерсти у крупного рогатого скота.
Кодоминированием называют такой тип взаимодействия аллельных генов, при котором оба аллеля проявляются, не ослабляя эффектов друг друга. Типичным примером кодоминирования является наследование групп крови по системе AB0.
Наследование групп крови по системе AB0
| Группа крови | Фенотип | Генотип | |
| агглютиногены | агглютинины | ||
| I (0) | — | $α$, $β$ | ii |
| II (A) | А | $β$ | IAIA или IAi |
| III (B) | В | $α$ | IBIB или IBi |
| IV (AB) | А и В | — | IAIB |
Как видно из таблицы, I, II и III группы крови наследуются по типу полного доминирования, тогда как IV (АВ) группа (генотип — IAIB) является случаем кодоминирования.
Сверхдоминирование — это явление, при котором в гетерозиготном состоянии доминантный признак проявляется намного сильнее, чем в гомозиготном; сверхдоминирование часто используется в селекции и считается причиной гетерозиса — явления гибридной силы.
Особым случаем взаимодействия аллельных генов можно считать так называемые летальные гены, которые в гомозиготном состоянии приводят к гибели организма чаще всего в эмбриональном периоде. Причиной гибели потомства является плейотропное действие генов серой окраски шерсти у каракулевых овец, платиновой окраски у лис и отсутствие чешуи у зеркальных карпов. При скрещивании двух гетерозиготных по этим генам особей расщепление по исследуемому признаку в потомстве будет равняться 2:1 вследствие гибели 1/4 потомства.
Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия. Комплементарность — это тип взаимодействия неаллельных генов, при котором для проявления определенного состояния признака необходимо присутствие как минимум двух доминантных аллелей разных пар. Например, у тыквы при скрещивании растений со сферическими (ААbb) и длинными (aaBB) плодами в первом поколении появляются растения с дисковидными плодами (AaBb).
К эпистазу относят такие явления взаимодействия неаллельных генов, при которых один неаллельный ген подавляет развитие признака другого. Например, у кур окраска оперения определяется одним доминантным геном, тогда как другой доминантный ген подавляет развитие окраски, в результате чего большинство кур имеет белое оперение.
Полимерией называют явление, при котором неаллельные гены оказывают одинаковое влияние на развитие признака. Таким образом чаще всего кодируются количественные признаки. Например, цвет кожи человека определяется как минимум четырьмя парами неаллельных генов — чем больше доминантных аллелей в генотипе, тем темнее кожа.
Генотип как целостная система
Генотип не является механической суммой генов, поскольку возможность проявления гена и форма его проявления зависят от условий среды. В данном случае под средой понимается не только окружающая среда, но и генотипическая среда — другие гены.
Проявление качественных признаков редко зависит от условий окружающей среды, хотя, если у горностаевого кролика выбрить участок тела с белой шерстью и прикладывать к нему пузырь со льдом, то со временем на этом месте вырастет черная шерсть.
Развитие количественных признаков намного сильнее зависит от условий окружающей среды. Например, если современные сорта пшеницы возделывать без применения минеральных удобрений, то ее урожайность будет существенно отличаться от генетически запрограммированных 100 и более центнеров с гектара.
Таким образом, в генотипе записаны лишь «способности» организма, однако проявляются они только во взаимодействии с условиями окружающей среды.
Кроме того, гены взаимодействуют друг с другом и, оказавшись в одном генотипе, могут сильно влиять на проявление действия соседних генов. Таким образом, для каждого отдельно взятого гена существует генотипическая среда. Возможно, что развитие любого признака связано с действием многих генов. Кроме того, выявлена зависимость нескольких признаков от одного гена. Например, у овса окраска цветочных чешуй и длина их ости определяются одним геном. У дрозофилы ген белой окраски глаза одновременно влияет на цвет тела и внутренних органов, длину крыльев, снижение плодовитости и уменьшение продолжительности жизни. Не исключено, что каждый ген одновременно является геном основного действия для «своего» признака и модификатором для других признаков. Таким образом, фенотип — это результат взаимодействия генов всего генотипа с окружающей средой в онтогенезе особи.
В связи с этим известный российский генетик М. Е. Лобашев определил генотип как систему взаимодействующих генов. Сложилась эта целостная система в процессе эволюции органического мира, при этом выживали лишь те организмы, у которых взаимодействие генов давало наиболее благоприятную реакцию в онтогенезе.
Генетика человека
Для человека как биологического вида в полной мере справедливы генетические закономерности наследственности и изменчивости, установленные для растений и животных. Вместе с тем генетика человека, изучающая закономерности наследственности и изменчивости у человека на всех уровнях его организации и существования, занимает особое место среди других разделов генетики.
Генетика человека одновременно является фундаментальной и прикладной наукой, поскольку занимается исследованием наследственных болезней человека, которых в настоящее время описано уже более 4 тыс. Она стимулирует развитие современных направлений общей и молекулярной генетики, молекулярной биологии и клинической медицины. В зависимости от проблематики генетика человека делится на несколько направлений, развившихся в самостоятельные науки: генетика нормальных признаков человека, медицинская генетика, генетика поведения и интеллекта, популяционная генетика человека. В связи с этим в наше время человек как генетический объект исследован едва ли не лучше, чем основные модельные объекты генетики: дрозофила, арабидопсис и др.
Биосоциальная природа человека накладывает значительный отпечаток на исследования в области его генетики вследствие позднего полового созревания и больших временных разрывов между поколениями, малочисленности потомства, невозможности направленных скрещиваний для генетического анализа, отсутствия чистых линий, недостаточной точности регистрации наследственных признаков и небольших родословных, невозможности создания одинаковых и строго контролируемых условий для развития потомков от разных браков, сравнительно большого числа плохо различающихся хромосом и невозможности экспериментального получения мутаций.
Методы изучения генетики человека
Методы, применяемые в генетике человека, принципиально не отличаются от общепринятых для других объектов — это генеалогический, близнецовый, цитогенетический, дерматоглифический, молекулярно-биологический и популяционно-статистический методы, метод гибридизации соматических клеток и метод моделирования. Их использование в генетике человека учитывает специфику человека как генетического объекта.
Близнецовый метод помогает определить вклад наследственности и влияние условий окружающей среды на проявление признака на основе анализа совпадения этих признаков у однояйцевых и разнояйцевых близнецов. Так, у большинства однояйцевых близнецов совпадают группы крови, цвет глаз и волос, а также целый ряд других признаков, тогда как корью болеют одновременно оба типа близнецов.
Дерматоглифический метод основан на исследовании индивидуальных особенностей кожных рисунков пальцев рук (дактилоскопия), ладоней и ступней ног. На основе этих особенностей он зачастую позволяет своевременно выявить наследственные заболевания, в частности хромосомные аномалии, такие как синдром Дауна, Шерешевского – Тернера и др.
Генеалогический метод — это метод составления родословных, с помощью которых определяют характер наследования изучаемых признаков, в том числе наследственных болезней, и прогнозируют рождение потомков с соответствующими признаками. Он позволил выявить наследственную природу таких заболеваний, как гемофилия, дальтонизм, хорея Гентингтона и др. еще до открытия основных закономерностей наследственности. При составлении родословных ведут записи о каждом из членов семьи и учитывают степень родства между ними. Далее на основании полученных данных с помощью специальной символики строится родословное древо.
Генеалогический метод можно использовать на одной семье, если есть сведения о достаточном количестве прямых родственников человека, родословная которого составляется — пробанда, — по отцовской и материнской линиям, в противном случае собирают сведения о нескольких семьях, в которых проявляется данный признак. Генеалогический метод позволяет установить не только наследуемость признака, но и характер наследования: доминантный или рецессивный, аутосомный или сцепленный с полом и т. д. Так, по портретам австрийских монархов Габсбургов было установлено наследование прогнатии (сильно выпяченной нижней губы) и «королевской гемофилии » у потомков британской королевы Виктории.
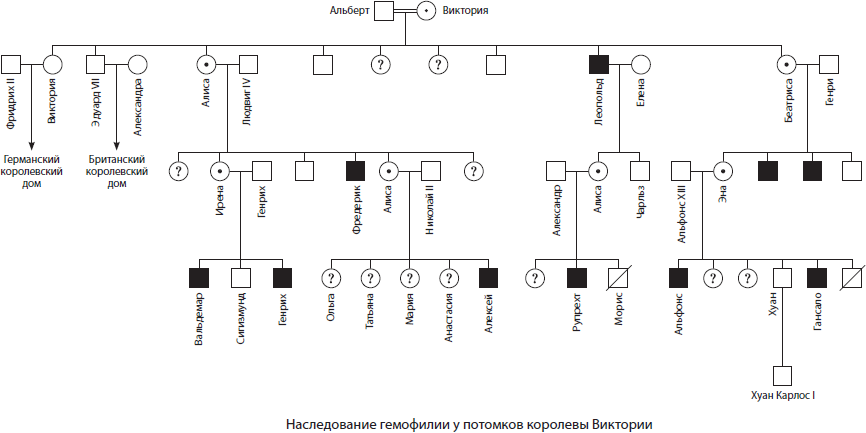
Решение генетических задач. Составление схем скрещивания
Все разнообразие генетических задач можно свести к трем типам:
- Расчетные задачи.
- Задачи на определение генотипа.
- Задачи на установление типа наследования признака.
Особенностью расчетных задач является наличие информации о наследовании признака и фенотипах родителей, по которым легко установить и генотипы родителей. В них требуется установить генотипы и фенотипы потомства.
Задача 1. Какую окраску будут иметь семена сорго, полученного в результате скрещивания чистых линий этого растения с темной и светлой окраской семян, если известно, что темная окраска доминирует над светлой? Какую окраску будут иметь семена растений, полученных от самоопыления этих гибридов?
Решение.
1. Обозначаем гены:
А — темная окраска семян, а — светлая окраска семян.
2. Составляем схему скрещивания:
а) сначала записываем генотипы родителей, которые по условию задачи являются гомозиготными:
$Р {♀АА}↙{\text"темные семена"}×{♂аа}↙{\text"светлые семена"}$
б) затем записываем гаметы в соответствии с правилом чистоты гамет:
Гаметы А a
в) попарно сливаем гаметы и записываем генотипы потомков:
F1 Аа
г) согласно закону доминирования все гибриды первого поколения будут иметь темную окраску, поэтому подписываем под генотипом фенотип.
Фенотип темные семена
3. Записываем схему следующего скрещивания:

Ответ: в первом поколении все растения будут иметь темную окраску семян, а во втором 3/4 растений будут иметь темные семена, а 1/4 — светлые.
Задача 2. У крыс черная окраска шерсти доминирует над бурой, а нормальная длина хвоста — над укороченным хвостом. Сколько потомков во втором поколении от скрещивания гомозиготных крыс с черной шерстью и нормальным хвостом с гомозиготными крысами с бурой шерстью и укороченным хвостом имели черную шерсть и укороченный хвост, если всего родилось 80 крысят?
Решение.
1. Записываем условие задачи:
А — черная шерсть, а — бурая шерсть;
В — нормальная длина хвоста, b — укороченный хвост.
F2 А_bb ?
2. Записываем схему скрещивания:

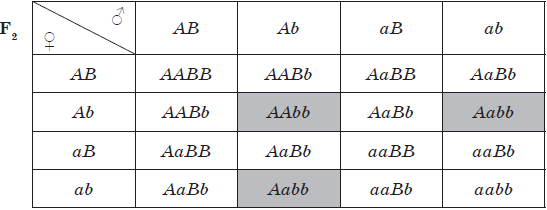
Примечание. Следует помнить, что буквенные обозначения генов записываются в алфавитном порядке, при этом в генотипах прописная буква всегда будет идти перед строчной: А — перед а, В — перед b и т. д.
Из решетки Пеннета следует, что доля крысят с черной шерстью и укороченным хвостом составляла 3/16.
3. Рассчитываем количество крысят с указанным фенотипом в потомстве второго поколения:
80 × 3/16 × 15.
Ответ: 15 крысят имели черную шерсть и укороченный хвост.
В задачах на определение генотипа также приводится характер наследования признака и ставится задание определить генотипы потомства по генотипам родителей или наоборот.
Задача 3. В семье, где отец имел ІІІ (В) группу крови по системе AB0, а мать — ІІ (А) группу, родился ребенок с І (0) группой крови. Определите генотипы родителей.
Решение.
1. Вспоминаем характер наследования групп крови:
Наследование групп крови по системе AB0
| Фенотип | Генотип |
| I (0) | ii |
| II (A) | IAIA или IAi |
| III (B) | IBIB или IBi |
| IV (AB) | IAIB |
2. Так как возможно по два варианта генотипов со II и III группами крови, схему скрещивания записываем следующим образом:
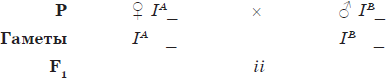
3. Из приведенной схемы скрещивания видим, что ребенок получил от каждого из родителей рецессивные аллели i, следовательно, родители были гетерозиготными по генам группы крови.
4. Дополняем схему скрещивания и проводим проверку наших предположений:
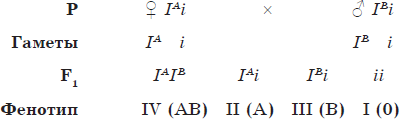
Таким образом, наши предположения подтвердились.
Ответ: родители гетерозиготны по генам групп крови: генотип матери — IAi, генотип отца — IВi.
Задача 4. Дальтонизм (цветовая слепота) наследуется как сцепленный с полом рецессивный признак. Какие дети могут родиться у мужчины и женщины, которые нормально различают цвета, хотя их родители были дальтониками, а матери и их родственники здоровы?
Решение.
1. Обозначаем гены:
ХD — нормальное цветовое зрение;
Хd — дальтонизм.
2. Устанавливаем генотипы мужчины и женщины, отцы которых были дальтониками.
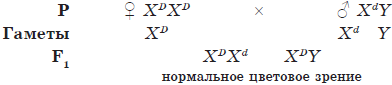
3. Записываем схему скрещивания для определения возможных генотипов детей:

Ответ: у всех девочек будет нормальное цветовое зрение (однако 1/2 девочек будет носителями гена дальтонизма), 1/2 мальчиков будет здорова, а 1/2 будет больна дальтонизмом.
В задачах на определение характера наследования признака приводятся только фенотипы родителей и потомства. Вопросами таких задач является именно выяснение характера наследования признака.
Задача 5. От скрещивания кур с короткими ногами было получено 240 цыплят, 161 из которых были коротконогими, а остальные — длинноногими. Как наследуется этот признак?
Решение.
1. Определяем расщепление в потомстве:
161 : 79 $≈$ 2 : 1.
Такое расщепление характерно для скрещиваний в случае летальных генов.
2. Так как кур с короткими ногами было вдвое больше, чем с длинными, допустим, что это доминантный признак, и именно этой аллели свойственен летальный эффект. Тогда исходные куры были гетерозиготными. Обозначаем гены:
С — короткие ноги, с — длинные ноги.
3. Записываем схему скрещивания:

Наши предположения подтвердились.
Ответ: коротконогость доминирует над длинноногостью, этой аллели свойствен летальный эффект.
Значение генетики для медицины. Наследственные болезни человека, их причины, профилактика. Вредное влияние мутагенов, алкоголя, наркотиков, никотина на генетический аппарат клетки. Защита среды от загрязнения мутагенами. Выявление источников мутагенов в окружающей среде (косвенно) и оценка возможных последствий их влияния на собственный организм
Значение генетики для медицины
Изучение закономерностей наследственности и изменчивости на растительных и животных объектах со временем привело к пониманию того, что наряду с инфекционными заболеваниями человека существует и значительное число (более 4000) наследственных болезней, развитие многих из которых обусловлено взаимодействием генетических программ и условий окружающей среды. Наследственные болезни затрагивают многие стороны обмена веществ, приводят к нарушениям структуры тканей и органов, а также психическим отклонениям.
Если ранее едва ли не единственными средствами профилактики наследственных заболеваний были запрет рожать детей женщинам, уже имеющим детей с отклонениями в развитии, а также принудительная стерилизация, то в настоящее время арсенал медицинских генетиков значительно расширился. Так, определение носительства родителями генов наследственных болезней и ранняя диагностика этих заболеваний еще до рождения ребенка позволяет избежать тяжелых последствий путем планирования семьи. Значительную помощь в этом отношении оказывает медико- генетическое консультирование.
Кроме того, профилактика многих заболеваний, в основе которой лежит исключение тех или иных веществ и продуктов из рациона питания, также позволяет предотвратить аномальное развитие и даже гибель больного.
Открытия в области молекулярной генетики позволили в последнее время совершить прорыв и в направлении исправления и замены патологических генов. Эта отрасль медицинской генетики называется генотерапией. Так, уже родился первый ребенок, у которого прямо в зиготе удалили ген наследственной формы рака.
Наследственные болезни человека, их причины, профилактика
Наследственные болезни составляют большую группу заболеваний, связанных с изменением генетической информации. По характеру их возникновения выделяют генные и хромосомные болезни, болезни несовместимости матери и плода, а также болезни с наследственной предрасположенностью.
Генные болезни обусловлены генными мутациями, возникшими как сотни лет назад и передающимися из поколения в поколение, так и недавно, в гаметах одного из родителей. Характерным примером такого рода заболеваний является «королевская» гемофилия, поразившая за столетие все монархии Европы. Несмотря на то, что в роду королевы Виктории не зафиксировано ни одного случая этой болезни, один из ее сыновей был болен, а две дочери стали носительницами гена. К генным заболеваниям относятся болезни обмена веществ (фенилкетонурия, глицинемия) и не связанные с ним аномалии (серповидноклеточная анемия, арахнодактилия). Вероятность проявления у детей генных заболеваний сильно повышается при близкородственных браках.
Хромосомные болезни обусловлены изменением структуры хромосом или их числа (хромосомные и геномные мутации), например, синдромы Дауна, Шерешевского – Тернера и Клайнфельтера. Синдром Дауна вызван наличием трех хромосом в 21-й паре. Он сопровождается не только задержкой в развитии ребенка, но и патологиями многих внутренних органов.
Синдром Шерешевского – Тернера (45, Х0), обусловленный недостатком одной половой хромосомы, негативно влияет на фенотип женщины с такой болезнью, которая, помимо низкого роста, может иметь патологии половой сферы и других органов.
Синдром Клайнфельтера связан с наличием лишней X-хромосомы в кариотипе (47, XXY). По фенотипу это высокий мужчина с телосложением по женскому типу, патологиями развития половых органов и их функционирования.
Рождение детей с вышеперечисленными хромосомными аномалиями может быть обусловлено вредными привычками, неблагоприятным состоянием окружающей среды и возрастом родителей.
Болезни несовместимости матери и плода являются следствием иммунологического конфликта, как, например, в случае резус-конфликта, который возникает, когда резус-отрицательная женщина вступает в брак с резус-положительным мужчиной, и у плода при этом резус будет положительным. Резус-конфликт может быть причиной гибели плода или рождения ребенка с серьезными патологиями нервной и других систем организма.
Болезни с наследственной предрасположенностью могут проявляться не сразу после рождения, а только через некоторое время, поскольку для активации соответствующих генов нужен целый комплекс факторов, в том числе характер питания и образ жизни человека. К таким заболеваниям относятся сахарный диабет и подагра.
Защита окружающей среды от загрязнения и ужесточение контроля за исполнением соответствующих решений, проверка поступающих на рынок продуктов питания, лекарственных препаратов и товаров бытового назначения на мутагенное и канцерогенное действие — необходимые меры по снижению частоты наследственных заболеваний у человека.
Не менее значимым для профилактики этой группы болезней является отсутствие у родителей вредных привычек, а также медико-генетическое консультирование супругов, в ходе которого можно не только определить вероятность рождения ребенка с отклонениями, но и скорректировать ход развития уже родившегося малыша.
Вредное влияние мутагенов, алкоголя, наркотиков, никотина на генетический аппарат клетки
Как физические, так и химические мутагены негативно влияют не только на саму материальную основу наследственности — ДНК, вызывая ее разрывы, перестройки и замещение одних нуклеотидов другими, но и подавляют активность ферментов репарации ДНК. Некоторые из этих веществ способны разрушать микротрубочки веретена деления и приводить к неправильному расхождению хромосом при митозе и мейозе. Вызванные таким образом мутации могут приводить к развитию у живущих особей раковых заболеваний, а у их потомков — наследственных заболеваний, таких как синдром Дауна.
Особую опасность для окружающих представляет курение, поскольку большая часть вредных веществ оседает не в легких курильщика, а в легких других людей, которых называют «пассивными курильщиками». Среди этих веществ содержатся многие канцерогенные (вызывающие рак) соединения, а также ионы тяжелых металлов.
Защита среды от загрязнения мутагенами
На современном этапе развития общества практически немыслим отказ от химической промышленности, интенсивных технологий в сельском хозяйстве, атомной энергетики и других источников мутагенов, однако степень загрязнения среды достигла такого критического уровня, что даже льды Антарктиды подвержены результатам хозяйственной деятельности человека. Это приводит не только к усилению процесса мутагенеза, но и к ухудшению состояния здоровья. Поэтому становятся все более актуальными вопросы очистки окружающей среды от загрязнения мутагенами. Для этого на государственном уровне устанавливаются предельные нормативы загрязнения вредными выбросами и отходами производства атмосферы, воды и почвы, а также значительные штрафные санкции за нарушение данных норм. В ряде стран мира вводятся даже ограничения на въезд в центр городов автомобилей и на курение в общественных местах, запрещается реклама продукции табачной и ликеро-водочной промышленности, проводятся интенсивные агитационные кампании по борьбе с курением и употреблением наркотиков.
Кроме того, вся продукция, поступающая на рынок, проверяется на соответствие установленным нормам содержания вредных веществ и изымается из продажи в случае нарушения данных норм. Постоянно контролируется и уровень загрязнения воды, атмосферы и почвы в крупных городах, а в случаях аварий проводится быстрая очистка окружающей среды от загрязнения с целью недопущения ухудшения условий жизни населения и нарушения экологического баланса. Например, после крушения в результате шторма десятка судов в Керченском проливе были собраны и откачаны более 1500 тонн мазута, которые уже вызвали гибель многих тысяч водоплавающих птиц и рыбы, но тем самым был предотвращен еще более существенный ущерб окружающей среде.
Выявление источников мутагенов в окружающей среде (косвенно) и оценка возможных последствий их влияния на организм человека
При всем разнообразии мутагенов существует только три основных пути их проникновения в тело человека: через дыхательные пути с воздухом, через пищеварительную систему с водой и пищей, а также через кожу. В связи с этим можно провести поиск источников мутагенов по тем средам, которые они загрязняют: воздух, вода и почва.
Загрязнение атмосферы осуществляется в основном автотранспортом и промышленными предприятиями. Его можно обнаружить по угнетению растительности вдоль автодорог, по быстро образующемуся слою пыли на окружающих предметах, по неприятному запаху. Определенную трудность в идентификации представляют некоторые источники ионизирующего и других видов излучения, например электромагнитного (станции мобильной связи, мобильные телефоны, мониторы телевизоров и компьютеров и т. д.), однако, например, в районе высоковольтных линий зачастую наблюдается либо угнетение, либо усиленный рост растений. Существенную угрозу жизни и здоровью окружающих представляют и курильщики, поэтому места, где они собираются, могут быть источниками химических мутагенов.
Выбросы загрязняющих веществ в водную среду могут приводить к изменению цвета воды, ее прозрачности, запаха и вкуса. Кроме того, в воде может наблюдаться усиленный рост водорослей («цветение» воды) с последующим замором рыбы и появлением гнилостного запаха.
Загрязнение почвы приводит чаще всего к тому, что нарушаются нормальные процессы роста и развития растений, появляются разнообразные уродства, меняется также видовой состав флоры. Например, всхожесть черной редьки на загрязненных территориях гораздо ниже, чем на незагрязненных. Отличается на таких территориях и видовой состав животного мира. Так, индикаторным видом является дождевой червь — он исчезает при увеличении степени загрязнения.
В зависимости от выявленных источников загрязнения определяют характер загрязнения и его степень, что может служить основанием для прогноза развития различных заболеваний (респираторных, аллергических, раковых и др.), а также мутагенного эффекта.
Селекция, ее задачи и практическое значение. Вклад Н. И. Вавилова в развитие селекции: учение о центрах многообразия и происхождения культурных растений; закон гомологических рядов в наследственной изменчивости. Методы селекции и их генетические основы. Методы выведения новых сортов растений, пород животных, штаммов микроорганизмов. Значение генетики для селекции. Биологические основы выращивания культурных растений и домашних животных
Селекция, ее задачи и практическое значение
Селекция (от лат. селектио — отбор) — это наука о методах создания пород домашних животных, сортов культурных растений и штаммов микроорганизмов с нужными человеку свойствами.
Под селекцией понимают также и сам процесс изменения живых организмов, осуществляемый человеком для своих потребностей. Современные селекционеры придерживаются точки зрения Н. И. Вавилова, согласно которой теоретической основой данной науки являются генетика и эволюционное учение.
Порода (сорт, штамм, чистая линия) — это популяция организмов, искусственно созданная человеком и характеризующаяся специфическим генофондом, наследственно закрепленными морфологическими и физиологическими признаками, определенным уровнем и характером продуктивности.
Задачей современной селекции является повышение продуктивности сортов растений и пород животных. Однако ныне важнейшими факторами интенсификации растениеводства и животноводства становится их перевод на промышленную основу, например, сортов овощей и фруктов, пригодных для машинной уборки, пород животных, предназначенных для содержания в животноводческих хозяйствах.
Достижения селекции растений, связанные с выведением высокопродуктивных сортов пшеницы, позволили осуществить так называемую «зеленую» революцию в середине ХХ века в Мексике, когда традиционные сорта были заменены новыми. Это позволило не только спасти от разорения мелкие фермерские хозяйства, но и решить продовольственную проблему в данном регионе. В целом с селекцией связывают надежды на преодоление дефицита продовольствия в мире, несмотря на глобальный экологический кризис, поразивший даже такие традиционно «хлебные» страны, как Испания и Аргентина.
Вклад Н. И. Вавилова в развитие селекции: учение о центрах многообразия и происхождения культурных растений; закон гомологических рядов в наследственной изменчивости
Первым этапом селекции было одомашнивание (доместикация), в процессе которого шел отбор по поведению животных и способности размножаться под контролем человека. Оно позволило сохранить огромное разнообразие признаков, в том числе неблагоприятных для вида. Всего человек окультурил около 150 видов растений и около 20 видов животных.
Выдающийся русский генетик и селекционер Н. И. Вавилов в ходе многочисленных экспедиций изучил мировые растительные ресурсы и установил, что наибольшее разнообразие форм определенного вида характерно для тех районов, где этот вид был введен в культуру. В соответствии с этим он определил семь центров происхождения культурных растений.
Центры происхождения культурных растений
| Название центра | Географическое положение | Примеры культурных растений |
| Южноазиатский тропический | Тропическая Индия, Индокитай, Южный Китай, о-ва Юго-Восточной Азии | Рис, сахарный тростник, цитрусовые, огурец, баклажан, черный перец и др. (50 % культурных растений) |
| Восточноазиатский | Центральный и Восточный Китай, Япония, Корея, Тайвань | Соя, просо, гречиха, плодовые и овощные культуры: слива, вишня, редька и др. (20 % культурных растений) |
| Юго-Западноазиатский | Малая Азия, Средняя Азия, Иран, Афганистан, Юго-Западная Индия | Пшеница, рожь, бобовые культуры, лен, конопля, репа, морковь, чеснок, виноград, абрикос, груша и др. (14 % культурных растений) |
| Средиземноморский | Страны Средиземноморского бассейна | Капуста, сахарная свекла, маслины, клевер, чечевица, кормовые травы (11 % культурных растений) |
| Абиссинский | Абиссинское нагорье Африки | Твердая пшеница, ячмень, кофейное дерево, сорго, бананы |
| Центральноамериканский | Южная Мексика | Кукуруза, длинноволокнистый хлопчатник, какао, тыква, табак |
| Южноамериканский | Южная Америка вдоль западного побережья | Картофель, ананас, хинное дерево |
Дальнейшие исследования позволили выделить уже 12 центров происхождения культурных растений, тесно связанных с центрами одомашнивания животных.
Н. И. Вавилов собрал также хранящуюся и поныне во Всероссийском институте растениеводства (г. Санкт-Петербург) мировую коллекцию культурных растений, которая и сейчас используется для выведения новых сортов и на основании изучения признаков культурных растений и близких к ним диких видов Н. И. Вавилов в 1920 году сформулировал закон гомологических рядов в наследственной изменчивости.
Закон гомологических рядов в наследственной изменчивости:
Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов.
Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство.
Данный закон позволил предположить наличие некоторых форм злаков, которые впоследствии были описаны, и, как позже было установлено, он является универсальным для всех живых организмов.
Генетической основой данного закона является то, что степень исторического родства прямо пропорциональна количеству их общих генов, вследствие чего и мутации этих генов могут быть сходными. В фенотипе это проявляется одинаковым характером изменчивости многих признаков у близких видов, родов и других таксонов.
Закон гомологических рядов наследственной изменчивости организмов объясняет направленность исторического развития родственных групп организмов. Опираясь на него и изучив наследственную изменчивость близких видов, в селекции планируют работу по созданию новых сортов растений и пород животных с определенным набором наследственных признаков. В систематике организмов этот закон позволяет предвидеть существование неизвестных науке систематических групп (видов, родов и т. д.) с подобными сочетаниями признаков, выявленных в близкородственных группах.
Методы селекции и их генетические основы
Основные методы селекции — гибридизация и искусственный отбор.
Гибридизация — это процесс образования или получения гибридов, в основе которого лежит объединение генетического материала разных клеток в одной клетке.
Для достижения результата в процессе гибридизации особое внимание уделяется подбору родительских пар. В селекции растений подбор ведется по определенным признакам с учетом генетической и географической удаленности; в селекции животных — только по хозяйственно ценным признакам, которые определяют по экстерьеру, родословной и потомству.
Выделяют родственную и неродственную гибридизации. Родственное скрещивание, или инбридинг, приводит к появлению чистых линий, но при этом снижается жизнеспособность потомства вследствие перехода различных летальных и полулетальных генов в гомозиготное состояние.
Неродственное скрещивание, или аутбридинг, бывает внутривидовым и межвидовым (в т. ч. отдаленная гибридизация). Аутбридинг в первом поколении дает эффект гетерозиса.
Гетерозис (от греч. гетерозис — изменение, перевоплощение) — явление повышения жизнеспособности и продуктивности у гибридов первого поколения по сравнению с исходными родительскими формами.
Данное явление объясняется благоприятным сочетанием родительских генов, а также переходом сублетальных и летальных аллелей в гетерозиготное состояние. Во втором и последующих поколениях эффект гетерозиса ослабевает вследствие расщепления генов и гомоготизации. У растений его эффект можно закрепить вегетативным или партеногенетическим размножением, удвоением числа хромосом и т. д. Эффект гетерозиса широко применяется в сельском хозяйстве, так как он позволяет существенно повысить урожайность растений (кукурузы, огурцов, томатов) и продуктивность животных (яйценоскость гибридов леггорнов и австралорнов, скорость роста и улучшение качества мяса бройлеров).
Несмотря на то, что с помощью отдаленной гибридизации уже созданы и успешно внедрены в сельскохозяйственное производство высокопродуктивные гибриды растений (пшенично-пырейный, пшеницы и ржи — тритикале, малины и ежевики), у животных (лошади и осла — мул, белуги и стерляди — бестер), основной проблемой данного метода является преодоление бесплодия гибридов. Бесплодие возникает в результате различий размеров, форм и количества хромосом в кариотипе родительских форм, вследствие чего хромосомы утрачивают способность конъюгировать в процессе мейоза. Преодолеть его можно за счет удвоения числа хромосом в кариотипе, и тогда хромосомы каждого из родителей будут конъюгировать с гомологичными им. Первым данный метод апробировал российский селекционер Г. Д. Карпеченко в процессе создания редечнокапустного гибрида с 36 хромосомами, тогда как у каждой из родительских форм их было по 18.
У животных решить проблему данным путем не представляется возможным вследствие увеличения дозы летальных аллелей, поэтому у них только в некоторых случаях один или оба пола плодовиты, как, например, самки гибридов яка с крупным рогатым скотом.
Искусственный отбор — процесс создания новых пород животных и сортов культурных растений путем систематического сохранения и размножения особей с определенными, ценными для человека признаками и свойствами в ряду поколений.
Выделяют две формы искусственного отбора: бессознательный, ведущийся без определенного плана, и методический, производимый с определенной целью. Примером искусственного отбора являются породы домашних голубей, выведенные от дикого скалистого голубя. Также он применяется в форме массового и индивидуального отбора. Массовый отбор является эффективным при высокой наследуемости признака. В основном он используется в селекции растений и микроорганизмов. При индивидуальном отборе учитываются не только показатели продуктивности или иные качества организма, но и наследование данного признака в ряду поколений. В комбинации с инбридингом он позволяет получить чистые линии. Индивидуальный отбор характерен для селекции животных и самоопыляющихся растений.
Теорию искусственного отбора создал великий английский ученый Ч. Дарвин. Основные положения своей теории он изложил в труде «Происхождение видов путем естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» и развил в дальнейшем в книге «Изменения домашних животных и культурных растений под влиянием одомашнивания».
Методы выведения новых сортов растений, пород животных, штаммов микроорганизмов
В связи с тем, что генетически запрограммированные резервы продуктивности культурных растений и животных уже практически исчерпаны, создание новых сортов и пород этих организмов требует кардинального изменения подхода к процессу селекции. В первую очередь перед началом селекционного процесса создается модель сорта или породы, которая учитывает современные требования к нему, после чего производится подбор методов, при помощи которых может быть достигнут искомый результат. Помимо описанных выше гибридизации и искусственного отбора, на современном этапе развития селекции широко используются также искусственный мутагенез, методы биотехнологии, клеточной и генной инженерии, клонирование.
Искусственным, или экспериментальным мутагенезом называют получение мутаций с помощью физических или химических агентов, например рентгеновского и ультрафиолетового излучения. Он позволяет получить как новые полезные генные мутации, так и геномные, в том числе добиться полиплоидизации. Однако далеко не все мутации происходят в ядерном геноме и способны передаваться в ряду поколений, поскольку в клетках животных имеются еще геномы митохондрий, а в клетках растений — митохондрий и пластид. Кроме того, мутации могут затронуть только соматические клетки, но не произойти в половых. В связи с этим многие мутантные формы растений размножаются только вегетативно.
В селекции растений широко применяются различные формы гибридизации и искусственного отбора. Однако гибриды довольно часто являются бесплодными, и поэтому их либо каждый раз выводят заново, либо размножают вегетативно. Для преодоления бесплодия гибридов у растений используется искусственный мутагенез, который позволяет получать полиплоидные сорта, отличающиеся более высокой урожайностью. С его помощью был получен ряд сортов сахарной свеклы, гречихи, редечно-капустный гибрид Г. Д. Карпеченко, а также новые высокоурожайные сорта ячменя и пшеницы, сорта растений с декоративными листьями.
В плодоводстве и декоративном цветоводстве невозможно в настоящее время обойтись без методов, разработанных и усовершенствованных одним из самых выдающихся российских селекционеров — И. В. Мичуриным, в особенности методов ментора, вегетативного сближения, посредника, смеси пыльцы и др. Например, метод ментора благодаря сочетанию свойств привоя и подвоя позволил ему вывести сорт груши бере зимняя.
Селекция животных использует те же методы, что и селекция растений, однако она учитывает биологические особенности этих организмов. Так, здесь на определенных стадиях селекционного процесса прибегают к инбридингу, однако весьма в ограниченных масштабах, поскольку это может привести к снижению жизнеспособности особей вследствие перевода летальных аллелей в гомозиготное состояние. Более широко распространенный в животноводстве аутбридинг может давать эффект гетерозиса, как в случае бройлеров — гибридов пород кур корниш и белого плимутрока, но при межвидовой гибридизации гибриды в основном бесплодны и их вегетативное размножение невозможно.
Еще одной трудностью селекционной работы в данной области является то, что у особей одного из полов могут не проявляться хозяйственно ценные признаки, например у петухов — яйценоскость, а у быков — молочность и жирность. В связи с этим от производителей получают «пробных » потомков, и только в том случае, если для последних характерны более высокие показатели исследуемого признака, производителей целесообразно использовать в дальнейшей работе. Для получения от них максимально возможного числа потомков применяют технологии искусственного осеменения, которые предусматривают получение и хранение половых клеток в течение длительного времени, а также искусственного оплодотворения «в пробирке» и пересадки в матку менее ценной в хозяйственном отношении самки — суррогатной матери.
Микроорганизмы в последнее время широко применяются в различных отраслях хозяйственной деятельности. Так, дрожжи используют в хлебопечении, виноделии, пивоварении и т. д. Другие грибы синтезируют в промышленных условиях антибиотики, лимонную кислоту и кормовые белки из отходов растениеводства и даже нефти. С помощью бактерий человек получает витамины, аминокислоты, инсулин, а также извлекает металлы из руд и промышленных отходов. Широко используются микроорганизмы в сельском и лесном хозяйстве для борьбы с вредителями.
Особенности организации и жизнедеятельности микроорганизмов не позволяют применять у них метод гибридизации, тогда как искусственный мутагенез с последующим отбором наиболее продуктивных штаммов дает прекрасные результаты. В некоторых случаях проводят искусственное скрещивание штаммов с помощью бактериофагов, способных переносить наследственную информацию из одной клетки бактерий в другую. Это позволило получить, например, высокопродуктивные штаммы грибов — продуцентов антибиотиков и витаминов.
Значение генетики для селекции
Хотя селекция и возникла как наука для удовлетворения практических потребностей человека, издавна применявшего гибридизацию особей с лучшими сочетаниями признаков для получения новых сортов растений и пород животных (именно на основе сравнения гибридов с родительскими формами начали формироваться основные представления о закономерностях наследования признаков), в настоящее время генетика является теоретической основой селекции. Опираясь на частную генетику различных объектов, селекционеры подбирают исходный материал для создания новых сортов растений, пород животных и штаммов микроорганизмов. При этом не только используются уже имеющиеся наследственные признаки, но и создаются новые благодаря применению метода искусственного мутагенеза, а также вносятся новые гены с помощью методов биотехнологии, не утрачивает своего значения и явление гетерозиса.
Окраска и структура меха пушных животных наследуются как качественные признаки, в связи с чем селекционеры используют их для выведения новых пород норки, лисицы, кролика и др. Продуктивность растений и крупного рогатого скота, напротив, являются количественными признаками, что также не может не учитываться в процессе выведения новых сортов и пород.
Значительную роль методы искусственного мутагенеза, клеточной и генной инженерии сыграли в выведении новых штаммов микроорганизмов, продуцирующих антибиотики, гормон роста человека, инсулин и др., а также в создании новых сортов растений и животных с измененными свойствами — генетически модифицированных организмов.
Биологические основы выращивания культурных растений и домашних животных
Для достижения генетически запрограммированной продуктивности сельскохозяйственные растения нуждаются в создании оптимальных условий. В первую очередь им, безусловно, необходима соответствующая интенсивность освещения, которая обеспечивает протекание процессов фотосинтеза, однако если пшеница требует высокой интенсивности света, то кофейные деревья необходимо выращивать в тени. Не менее существенным фактором является и достаточное количество влаги в почве, что можно обеспечить в основном благодаря созданию оросительных систем, хотя в настоящее время все чаще прибегают к капельному поливу. Еще одним важным условием повышения урожайности сельскохозяйственных культур является обеспечение их элементами минерального питания. Эту проблему частично можно решить путем внесения в почву удобрений, что, однако, сопряжено с риском чрезмерного их накопления и смыва в близлежащие водоемы. Поэтому стараются применять многопольные севообороты, в которые включают бобовые, образующие симбиоз с клубеньковыми бактериями, переводящими атмосферный азот в доступную для растений форму.
С момента зарождения земледелия культурные растения страдают от вредителей и возбудителей различных заболеваний, которые снижают их урожайность, а в некоторых случаях и полностью уничтожают посевы. Причиной таких стихийных бедствий является их пониженная устойчивость к факторам среды и занятие больших площадей одним видом растений. Для борьбы с вредителями растений долгое время использовали химические вещества — пестициды, однако со временем выяснилось, что появились новые расы, устойчивые к этим веществам, а сами пестициды обладают токсическим и мутагенным действием. Поэтому в настоящее время во многих странах использование пестицидов существенно ограничено или вовсе запрещено. В связи с этим на передний план выходят биологические методы борьбы с вредителями, которые связаны либо с массовым размножением хищника или паразита данного вредителя, либо с нарушением размножения вредителя путем отлова самцов, а также с искусственной стерилизацией самцов, которые не дают потомков в результате скрещивания с нормальными самками.
Сельскохозяйственные животные, выращиваемые по интенсивным технологиям, также нуждаются в особых условиях. В первую очередь, им требуются сбалансированные корма, в которые ранее добавляли белок, полученный в результате бактериального синтеза, однако затем от него отказались, поскольку он мог вызывать аллергии не только у животных, но и у работников предприятий и жителей близлежащих населенных пунктов. Поэтому в настоящее время корма составляются большей частью на растительной основе.
Перспективы развития растениеводства и животноводства и, в конечном итоге, решение проблемы кризиса продовольствия связаны в основном с прогрессом биотехнологии, клеточной и генной инженерии.
Биотехнология, ее направления. Клеточная и генная инженерия, клонирование. Роль клеточной теории в становлении и развитии биотехнологии. Значение биотехнологии для развития селекции, сельского хозяйства, микробиологической промышленности, сохранения генофонда планеты. Этические аспекты развития некоторых исследований в биотехнологии (клонирование человека, направленные изменения генома)
Биотехнология, ее направления
Биотехнология — это применение биологических процессов и использование живых организмов в промышленности, медицине, сельском хозяйстве и других отраслях человеческой деятельности.
Несмотря на то, что биологические процессы издавна используются человеком в хлебопечении, сыроварении, виноделии, пивоварении, научный этап развития биотехнологии начался с 70-х годов ХIХ века с открытием Л. Пастером процесса брожения, а столетием позже биотехнология превратилась в бурно развивающуюся отрасль. В настоящее время прогресс в области биотехнологии тесно связан с применением методов генной и клеточной инженерии, а также клонированием.
В качестве основных направлений биотехнологии рассматриваются получение продуктов питания, кормовых добавок и ценных кормовых белков, лекарственных препаратов и средств диагностики, биотоплива, борьба с загрязнением окружающей среды, защита растений от вредителей и болезней, а также создание штаммов микроорганизмов, сортов растений и пород животных с новыми полезными свойствами.
В настоящее время в хлебопекарной и кондитерской промышленностях, пивоварении и виноделии применяются различные штаммы дрожжей. Благодаря способности осуществлять спиртовое брожение для них нашлось место и в технологиях выработки биотоплива, например, биодизеля из растительного сырья, особенно рапса. Другие микроскопические грибы широко используют для получения кефира, сыров, антибиотиков, лимонной кислоты, кормовых белков и т. д.
Без бактерий невозможно получить никакие кисломолочные продукты, в том числе кефир, йогурт и сыры. Брожение, осуществляемое молочнокислыми бактериями, используется и в процессах приготовления квашеных овощей, а также силоса, поскольку накапливающиеся при этом продукты реакции угнетают развитие других микроорганизмов. Не меньшую роль бактерии играют и в фармацевтической промышленности, где они культивируются с целью получения витаминов, гормонов и ферментов. Первой микробиологический синтез гормона инсулина с помощью методов генной инженерии «освоила» кишечная палочка Escherichia coli.
Очистка окружающей среды ведется в основном в двух направлениях: разложение органических остатков и накопление отдельных химических элементов, органических и неорганических веществ некоторыми видами бактерий, водорослей и простейших. С помощью методов селекции и генной инженерии уже выведены штаммы бактерий, способные разлагать соединения, утилизировать которые встречающиеся в природе виды неспособны, например пластмассы и полиэтилен. В процессе расщепления органических остатков бактерии могут выделять и горючие газы, в том числе метан, что легло в основу технологий получения биогаза из отходов растениеводства и животноводства.
В связи с тем, что бактерии, грибы и вирусы способны эффективно бороться с вредителями сельского и лесного хозяйства, а также с возбудителями и переносчиками заболеваний, их штаммы используют для приготовления биопрепаратов. Преимущество этих биологических методов борьбы состоит в том, что они не только снижают численность паразитов, будучи безвредными для других организмов, но и не загрязняют при этом окружающую среду токсичными соединениями.
Клеточная и генная инженерия, клонирование
Клеточная инженерия — метод конструирования клеток нового типа на основе их культивирования на питательной среде, гибридизации и реконструкции. При этом в клетки вводят новые хромосомы, ядра и другие клеточные структуры.
Достижения клеточной инженерии растений, которая позволяет сформировать целое растение, в том числе с измененными свойствами, из отдельной клетки, нашли широкое применение в растениеводстве и селекции. Так, стали возможными соматическая гибридизация, клеточная селекция, гаплоидизация, преодоление нескрещиваемости в культуре и другие приемы.
Технологии искусственного оплодотворения, за разработку которых присуждена Нобелевская премия в области физиологии и медицины в 2010 году, также базируются на методах клеточной инженерии.
Генная инженерия — это отрасль молекулярной биологии и генетики, задачей которой является конструирование генетических структур по заранее намеченному плану, создание организмов с новой генетической программой. Во многих случаях это сводится к переносу необходимых генов от одного вида живых организмов к другому, зачастую очень далекому по происхождению.
Переносу генов предшествует кропотливая работа по выявлению нужного гена в геноме организма- донора (вируса, бактерии, растения, животного, гриба) и его выделению. Это наиболее трудная часть работы, поскольку вместе со структурным геном необходимо перенести и регуляторные. Затем необходимо встроить данный участок молекулы ДНК в генетический вектор (переносчик ДНК). В качестве векторов чаще всего используют вирусы, плазмиды бактерий, хромосомы митохондрий и пластид, а также искусственно сконструированные молекулы ДНК. Процесс введения вектора новой ДНК в клетку-хозяина называется трансформацией. Последний этап работы заключается в размножении организмов-хозяев и отборе тех из них, в которых «прижился» введенный ген. В настоящее время применяют и прямое введение ДНК в клетки эукариот с помощью электрических разрядов, генной пушки и другими способами. Полученные в результате переноса генов организмы называются генетически модифицированными, или трансгенными.
Клонирование — это получение многочисленных копий гена, белка, клетки или организма. Клонирование генов чаще всего осуществляется с помощью бактерий и вирусов, поскольку, например, одна вирусная частица бактериофага, в которой содержится нужный ген, за один день может образовать более 1012 идентичных копий себя и этой молекулы.
Клонирование растений также не представляет значительной трудности, поскольку клетки растений тотипотентны, т. е. из одной клетки можно восстановить целый организм, особенно если культивировать эти клетки на питательной среде со всеми необходимыми веществами.
Массовое размножение генетически идентичных животных долгое время сталкивалось с таким существенным препятствием, как отсутствие способности к бесполому размножению у высших животных. Однако в 1997 году эта проблема была разрешена с получением первого клонированного организма — овцы Долли. Для клонирования были взяты клетки молочной железы ее генетической матери, а также яйцеклетки суррогатной матери. Ядра яйцеклеток удалялись, а на их место вводились ядра клеток молочной железы. После стимуляции развития зиготы электрическим током делящийся зародыш короткий промежуток времени культивировали на питательной среде, а затем вводили в матку суррогатной матери. К сожалению, из пяти пересаженных эмбрионов выжил лишь один.
В настоящее время клонирован уже целый ряд видов животных — мыши, собаки, коровы и др., однако клонирование человека запрещено законодательством многих государств и международными договорами.
Заманчивые перспективы перед человечеством раскрываются в области терапевтического клонирования — воспроизведения отдельных органов. Так, в настоящее время широко используются клонированная кожа, клетки соединительной ткани и другие части организма.
Роль клеточной теории в становлении и развитии биотехнологии
Создание клеточной теории позволило связать наследственность и изменчивость с их материальной основой — ДНК, а также определить, что клетка является единицей строения, жизнедеятельности и развития живых организмов. Поэтому дальнейшее внимание исследователей в области биотехнологии было сосредоточено именно на клетке как основном объекте. Уже в середине ХХ века были получены первые растения, выращенные из отдельных клеток на питательной среде, а в 1973 году родился первый «ребенок из пробирки». Операции с клетками (генная и клеточная инженерии) позволили клонировать сначала холоднокровных животных, а затем и млекопитающих.
Значение биотехнологии для развития селекции, сельского хозяйства, микробиологической промышленности, сохранения генофонда планеты
Прогресс биотехнологии позволил совершить прорыв в таких отраслях человеческой деятельности, как селекция, сельское хозяйство, медицина, фармация и др., поскольку появились возможности не только для изменения свойств организмов, но и для ускорения процесса их создания. Так, введение в растения бактериальных генов устойчивости к поеданию насекомыми и поражению вирусами, а также способных расти на бедных или загрязненных почвах способствует решению продовольственной проблемы, особенно в странах с быстро растущим населением. В настоящее время значительная часть посевных площадей занята трансгенными культурами в США, Канаде и Китае.
Кроме того, культивирование клеток растений на фоне высоких концентраций солей и других соединений позволяет сократить сроки выведения новых сортов пшеницы, сои и других важнейших сельскохозяйственных культур до одного-двух лет.
Клонирование животных, особенно с генетически измененными признаками и свойствами, позволяет вывести более продуктивные породы и добиться их быстрого размножения, однако этот процесс пока еще слишком трудоемок и дорог, чтобы применяться в промышленном масштабе.
Трансформация бактерий позволила уже в начале 80-х годов ХХ века получать биологически активные вещества — инсулин, соматотропный гормон, интерферон, которые применяются в медицине, а также создать новые штаммы микроорганизмов, предназначенных для очистки сточных вод, ликвидации нефтяных разливов и т. д. Путем селекции выведены также и формы бактерий, с помощью которых получают антибиотики, извлекают цветные металлы, получают биогаз.
В будущем возможно использование клонирования в сочетании с другими отраслями биотехнологии не только для размножения растений, микроорганизмов и грибов, но и для восстановления исчезнувших видов животных, возобновления природных популяций исчезающих видов. Однако для этого необходимо вначале создать генные банки, поскольку ДНК довольно быстро подвергается разрушению в окружающей среде.
Этические аспекты развития некоторых исследований в биотехнологии (клонирование человека, направленные изменения генома)
Расширение сферы влияния биотехнологии, с одной стороны, преследует благородные цели, поскольку с ее помощью стало возможным преодоление бесплодия, лечение многих наследственных и приобретенных заболеваний, а также решение продовольственных и экологических проблем современности. С другой стороны, активное вторжение современных технологий в медицину не может не настораживать, поскольку это сопряжено с операциями с клетками и тканями человека. Например, не совсем ясно, почему по американским законам при искусственном оплодотворении берется две донорские яйцеклетки, но пересаживается только одна из них, тогда как вторая замораживается, помещается в специальный банк и не выдается родителям даже по специальному запросу.
Большинство стран законодательно ограничило эксперименты по клонированию человека в основном по этическим соображениям, поскольку они направлены не просто на воспроизведение человека, но и на последующее использование клеток, тканей и органов зародыша для экспериментов, а также в качестве их донора. В связи с этим во всем мире активно обсуждается вопрос о допустимости подобных действий.
Применение генных технологий в создании новых сортов растений, пород животных и штаммов микроорганизмов также вызывает некоторые опасения, поскольку их попадание в окружающую среду может вызвать неконтролируемое распространение, например, раковых генов, и привести к необратимым последствиям для жизни и здоровья человека. Так, опыление пыльцой трансгенных растений генетически немодифицированных сортов и видов может стимулировать появление сверхустойчивых к химическим и биологическим средствам борьбы сорняков.
Особую опасность представляет внесение новых генов в сбалансированный геном, откуда они могут быть исключены в любой момент, что может привести к появлению каких-либо вирусоподобных организмов.
Потребление продуктов, полученных с использованием генетически модифицированных организмов, по некоторым данным, приводит к существенным нарушениям в репродуктивной сфере человека, а в перспективе может угрожать и самой жизни, поскольку мутировавший лишь по одному нуклеотиду ген устойчивости картофеля к поеданию колорадским жуком кодирует белок, смертельно опасный уже и для человека. И хотя это является маловероятным, поскольку ДНК потребляемых нами продуктов должна расщепляться в кишечнике, все же такая вероятность существует, и сбрасывать ее со счетов не приходится.
Сравнительно слабая изученность проблем клонирования и применения генных технологий заставляет многие правительства принимать решения по ограничению сферы их применения и специальной маркировке продуктов питания, полученных таким способом, с целью информирования.

Онлайн-школа «Турбо»
- Прямая связь с преподавателем
- Письменные дз с проверкой
- Интересные онлайн-занятия
- Душевное комьюнити