Клетка как биологическая система (соответствие)

Строение клетки. Взаимосвязь строения и функций частей и органоидов клетки — основа ее целостности
Строение клетки
Строение прокариотических и эукариотических клеток
Основными структурными компонентами клеток являются плазматическая мембрана, цитоплазма и наследственный аппарат. В зависимости от особенностей организации различают два основных типа клеток: прокариотические и эукариотические. Главным отличием прокариотических клеток от эукариотических является организация их наследственного аппарата: у прокариот он находится непосредственно в цитоплазме (эта область цитоплазмы называется нуклеоидом) и не отделен от нее мембранными структурами, тогда как у эукариот бульшая часть ДНК сосредоточена в ядре, окруженном двойной мембраной. Кроме того, генетическая информация прокариотических клеток, находящаяся в нуклеоиде, записана в кольцевой молекуле ДНК, а у эукариот молекулы ДНК незамкнутые.
В отличие от эукариот, цитоплазма прокариотических клеток содержит также небольшое количество органоидов, тогда как для эукариотических характерно значительное разнообразие этих структур.
Строение и функции биологических мембран
Строение биомембраны. Мембраны, ограничивающие клетки и мембранные органоиды эукариотических клеток, имеют общий химический состав и строение. В их состав входят липиды, белки и углеводы. Липиды мембраны представлены в основном фосфолипидами и холестерином. Большинство белков мембран относится к сложным белкам, например гликопротеинам. Углеводы не встречаются в мембране самостоятельно, они связаны с белками и липидами. Толщина мембран составляет 7–10 нм.
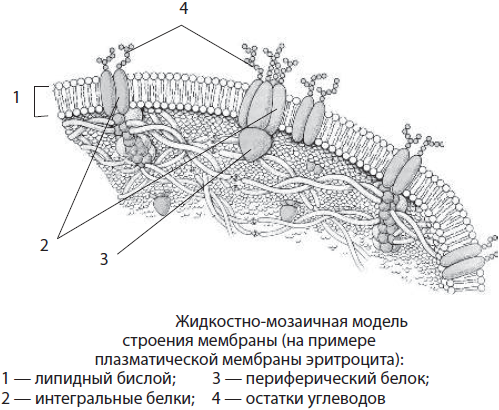
Согласно общепринятой в настоящее время жидкостно-мозаичной модели строения мембран, липиды образуют двойной слой, или липидный бислой, в котором гидрофильные «головки» молекул липидов обращены наружу, а гидрофобные «хвосты» спрятаны вовнутрь мембраны. Эти «хвосты» благодаря своей гидрофобности обеспечивают разделение водных фаз внутренней среды клетки и ее окружения. С липидами с помощью различных типов взаимодействия связаны белки. Часть белков расположена на поверхности мембраны. Такие белки называют периферическими, или поверхностными. Другие белки частично или полностью погружены в мембрану — это интегральные, или погруженные белки. Белки мембран выполняют структурную, транспортную, каталитическую, рецепторную и другие функции.
Мембраны не похожи на кристаллы, их компоненты постоянно находятся в движении, вследствие чего между молекулами липидов возникают разрывы — поры, через которые в клетку могут попадать или покидать ее различные вещества.
Биологические мембраны различаются по расположению в клетке, химическому составу и выполняемым функциям. Основные типы мембран — плазматическая и внутренние. Плазматическая мембрана содержит около 45 % липидов (в т. ч. гликолипидов), 50 % белков и 5 % углеводов. Цепочки углеводов, входящих в состав сложных белков-гликопротеинов и сложных липидов-гликолипидов, выступают над поверхностью мембраны. Гликопротеины плазмалеммы чрезвычайно специфичны. Так, например, по ним происходит взаимное узнавание клеток, в том числе сперматозоида и яйцеклетки.
На поверхности животных клеток углеводные цепочки образуют тонкий поверхностный слой — гликокаликс. Он выявлен почти во всех животных клетках, но степень его выраженности неодинакова (10–50 мкм). Гликокаликс обеспечивает непосредственную связь клетки с внешней средой, в нем происходит внеклеточное пищеварение; в гликокаликсе размещены рецепторы. Клетки бактерий, растений и грибов, помимо плазмалеммы, окружены еще и клеточными оболочками.
Внутренние мембраны эукариотических клеток разграничивают различные части клетки, образуя своеобразные «отсеки» — компартменты, что способствует разделению различных процессов обмена веществ и энергии. Они могут различаться по химическому составу и выполняемым функциям, но общий план строения у них сохраняется.
Функции мембран:
- Ограничивающая. Заключается в том, что они отделяют внутреннее пространство клетки от внешней среды. Мембрана является полупроницаемой, то есть ее свободно преодолевают только те вещества, которые необходимы клетке, при этом существуют механизмы транспорта необходимых веществ.
- Рецепторная. Связана в первую очередь с восприятием сигналов окружающей среды и передачей этой информации внутрь клетки. За эту функцию отвечают специальные белки-рецепторы. Мембранные белки отвечают еще и за клеточное узнавание по принципу «свой-чужой», а также за образование межклеточных соединений, наиболее изученными из которых являются синапсы нервных клеток.
- Каталитическая. На мембранах расположены многочисленные ферментные комплексы, вследствие чего на них происходят интенсивные синтетические процессы.
- Энерготрансформирующая. Связана с образованием энергии, ее запасанием в виде АТФ и расходованием.
- Компартментализация. Мембраны разграничивают также пространство внутри клетки, разделяя тем самым исходные вещества реакции и ферменты, которые могут осуществлять соответствующие реакции.
- Образование межклеточных контактов. Несмотря на то, что толщина мембраны настолько мала, что ее невозможно различить невооруженным глазом, она, с одной стороны, служит достаточно надежным барьером для ионов и молекул, в особенности водорастворимых, а с другой — обеспечивает их перенос в клетку и наружу.
- Транспортная.
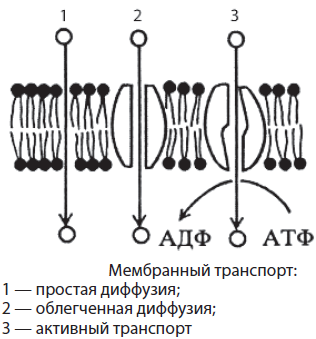
Мембранный транспорт. В связи с тем, что клетки как элементарные биологические системы являются открытыми системами, для обеспечения обмена веществ и энергии, поддержания гомеостаза, роста, раздражимости и других процессов требуется перенос веществ через мембрану — мембранный транспорт. В настоящее время транспорт веществ через мембрану клетки делят на активный, пассивный, эндо- и экзоцитоз.
Пассивный транспорт — это вид транспорта, который происходит без затраты энергии от большей концентрации к меньшей. Растворимые в липидах небольшие неполярные молекулы (О2, СО2) легко проникают в клетку путем простой диффузии. Нерастворимые же в липидах, в том числе заряженные небольшие частицы, подхватываются белкамипереносчиками или проходят через специальные каналы (глюкоза, аминокислоты, К+, PO43-). Такой вид пассивного транспорта называется облегченной диффузией. Вода поступает в клетку через поры в липидной фазе, а также по специальным каналам, выстланным белками. Транспорт воды через мембрану называется осмосом.
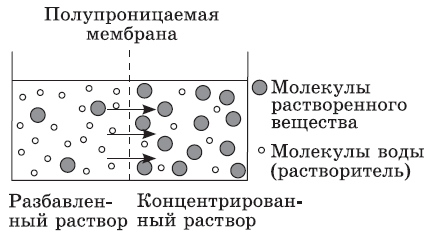
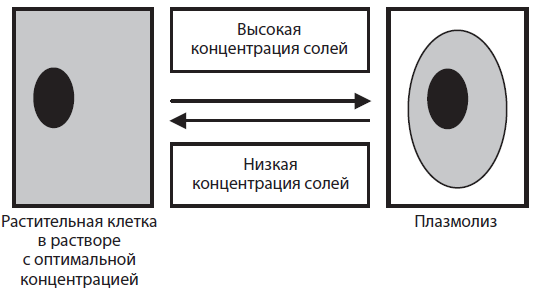
Осмос имеет чрезвычайно важное значение в жизни клетки, так как если ее поместить в раствор с более высокой концентрацией солей, чем в клеточном растворе, то вода начнет выходить из клетки, и объем живого содержимого начнет уменьшаться. У животных клеток происходит съеживание клетки в целом, а у растительных — отставание цитоплазмы от клеточной стенки, которое называется плазмолизом. При помещении клетки в менее концентрированный, чем цитоплазма, раствор, транспорт воды происходит в обратном направлении — в клетку. Однако существуют пределы растяжимости цитоплазматической мембраны, и животная клетка в конце концов разрывается, а у растительной этого не позволяет сделать прочная клеточная стенка. Явление заполнения клеточным содержимым всего внутреннего пространства клетки называется деплазмолизом. Внутриклеточную концентрацию солей следует учитывать при приготовлении лекарственных препаратов, особенно для внутривенного введения, так как это может приводить к повреждению клеток крови (для этого используют физиологический раствор с концентрацией 0,9 % хлорида натрия). Это не менее важно при культивировании клеток и тканей, а также органов животных и растений.
Активный транспорт протекает с затратой энергии АТФ от меньшей концентрации вещества к большей. Он осуществляется с помощью специальных белков-насосов. Белки перекачивают через мембрану ионы К+, Na+, Са2+ и другие, что способствует транспорту важнейших органических веществ, а также возникновению нервных импульсов и т. д.
Эндоцитоз — это активный процесс поглощения веществ клеткой, при котором мембрана образует впячивания, а затем формирует мембранные пузырьки — фагосомы, в которых заключены поглощаемые объекты. Затем с фагосомой сливается первичная лизосома, и образуется вторичная лизосома, или фаголизосома, или пищеварительная вакуоль. Содержимое пузырька расщепляется ферментами лизосом, а продукты расщепления поглощаются и усваиваются клеткой. Непереваренные остатки удаляются из клетки путем экзоцитоза. Различают два основных вида эндоцитоза: фагоцитоз и пиноцитоз.
Фагоцитоз — это процесс захвата клеточной поверхностью и поглощения клеткой твердых частиц, а пиноцитоз — жидкости. Фагоцитоз протекает в основном в животных клетках (одноклеточные животные, лейкоциты человека), он обеспечивает их питание, а часто и защиту организма . Путем пиноцитоза происходит поглощение белков, комплексов антиген-антитела в процессе иммунных реакций и т. д. Однако путем пиноцитоза или фагоцитоза в клетку также попадают многие вирусы. В клетках растений и грибов фагоцитоз практически невозможен, так как они окружены прочными клеточными оболочками.
Экзоцитоз — процесс, обратный эндоцитозу. Таким образом выделяются непереваренные остатки пищи из пищеварительных вакуолей, выводятся необходимые для жизнедеятельности клетки и организма в целом вещества. Например, передача нервных импульсов происходит благодаря выделению посылающим импульс нейроном химических посредников — медиаторов, а в растительных клетках так выделяются вспомогательные углеводы клеточной оболочки.
Клеточные оболочки клеток растений, грибов и бактерий. Снаружи от мембраны клетка может выделять прочный каркас — клеточную оболочку, или клеточную стенку.
У растений основу клеточной оболочки составляет целлюлоза, упакованная в пучки по 50–100 молекул. Промежутки между ними заполняют вода и другие углеводы. Оболочка растительной клетки пронизана канальцами — плазмодесмами, через которые проходят мембраны эндоплазматической сети. По плазмодесмам осуществляется транспорт веществ между клетками. Однако транспорт веществ, например воды, может происходить и по самим клеточным стенкам. Со временем в клеточной оболочке растений накапливаются различные вещества, в том числе дубильные или жироподобные, что приводит к одревеснению или опробковению самой клеточной стенки, вытеснению воды и отмиранию клеточного содержимого. Между клеточными стенками соседних клеток растений располагаются желеобразные прокладки — срединные пластинки, которые скрепляют их между собой и цементируют тело растения в целом. Они разрушаются только в процессе созревания плодов и при опадании листьев.
Клеточные стенки клеток грибов образованы хитином — углеводом, содержащим азот. Они достаточно прочны и являются внешним скелетом клетки, но все же, как и у растений, препятствуют фагоцитозу.
У бактерий в состав клеточной стенки входит углевод с фрагментами пептидов — муреин, однако его содержание существенно различается у разных групп бактерий. Поверх от клеточной стенки могут выделяться также иные полисахариды, образующие слизистую капсулу, защищающую бактерии от внешних воздействий.
Оболочка определяет форму клетки, служит механической опорой, выполняет защитную функцию, обеспечивает осмотические свойства клетки, ограничивая растяжение живого содержимого и предотвращая разрыв клетки, увеличивающейся вследствие поступления воды. Кроме того, клеточную стенку преодолевают вода и растворенные в ней вещества, прежде чем попасть в цитоплазму или, наоборот, при выходе из нее, при этом по клеточным стенкам вода транспортируется быстрее, чем по цитоплазме.
Цитоплазма
Цитоплазма — это внутреннее содержимое клетки. В нее погружены все органоиды клетки, ядро и разнообразные продукты жизнедеятельности.
Цитоплазма связывает все части клетки между собой, в ней протекают многочисленные реакции обмена веществ. Цитоплазма отделяется от окружающей среды и делится на отсеки мембранами, то есть клеткам присуще мембранное строение. Она может находиться в двух состояниях — золя и геля. Золь — это полужидкое, киселеобразное состояние цитоплазмы, при котором процессы жизнедеятельности протекают наиболее интенсивно, а гель — более плотное, студнеобразное состояние, затрудняющее протекание химических реакций и транспорт веществ.
Жидкая часть цитоплазмы без органоидов называется гиалоплазмой. Гиалоплазма, или цитозоль, представляет собой коллоидный раствор, в котором находится своеобразная взвесь достаточно крупных частиц, например белков, окруженных диполями молекул воды. Осаждения этой взвеси не происходит вследствие того, что они имеют одинаковый заряд и отталкиваются друг от друга.
Органоиды
Органоиды — это постоянные компоненты клетки, выполняющие определенные функции.
В зависимости от особенностей строения их делят на мембранные и немембранные. Мембранные органоиды, в свою очередь, относят к одномембранным (эндоплазматическая сеть, комплекс Гольджи и лизосомы) или двумембранным (митохондрии, пластиды и ядро). Немембранными органоидами являются рибосомы, микротрубочки, микрофиламенты и клеточный центр. Прокариотам из перечисленных органоидов присущи только рибосомы.
Строение и функции ядра. Ядро — крупный двумембранный органоид, лежащий в центре клетки или на ее периферии. Размеры ядра могут колебаться в пределах 3–35 мкм. Форма ядра чаще сферическая или эллипсоидная, однако имеются также палочковидные, веретеновидные, бобовидные, лопастные и даже сегментированные ядра. Некоторые исследователи считают, что форма ядра соответствует форме самой клетки.
Большинство клеток имеет одно ядро, но, например, в клетках печени и сердца их может быть два, а в ряде нейронов — до 15. Волокна скелетных мышц содержат обычно много ядер, однако они не являются клетками в полном смысле этого слова, поскольку образуются в результате слияния нескольких клеток.
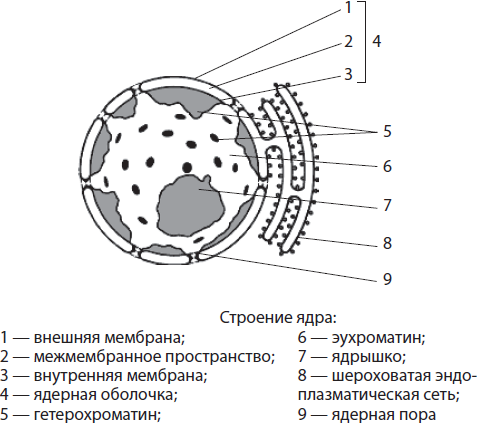
Ядро окружено ядерной оболочкой, а его внутреннее пространство заполнено ядерным соком, или нуклеоплазмой (кариоплазмой), в которую погружены хроматин и ядрышко. Ядро выполняет такие важнейшие функции, как хранение и передача наследственной информации, а также контроль жизнедеятельности клетки.
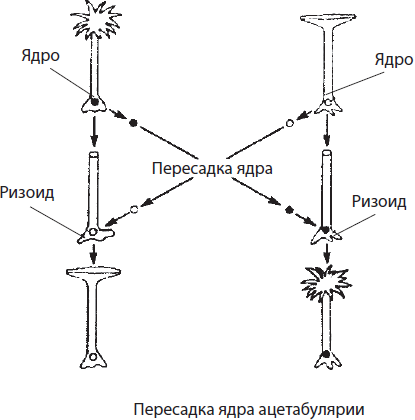
Роль ядра в передаче наследственной информации была убедительно доказана в экспериментах с зеленой водорослью ацетабулярией. В единственной гигантской клетке, достигающей в длину 5 см, различают шляпку, ножку и ризоид. При этом она содержит только одно ядро, расположенное в ризоиде. В 1930-е годы И. Хеммерлинг пересадил ядро одного вида ацетабулярии с зеленой окраской в ризоид другого вида, с коричневой окраской, у которого ядро было удалено. Через некоторое время у растения с пересаженным ядром выросла новая шляпка, как у водоросли- донора ядра. В то же время отделенные от ризоида шляпка или ножка, не содержащие ядра, через некоторое время погибали.
Ядерная оболочка образована двумя мембранами — наружной и внутренней, между которыми есть пространство. Межмембранное пространство сообщается с полостью шероховатой эндоплазматической сети, а наружная мембрана ядра может нести рибосомы. Ядерная оболочка пронизана многочисленными порами, окантованными специальными белками. Через поры происходит транспорт веществ: в ядро попадают необходимые белки (в т. ч. ферменты), ионы, нуклеотиды и другие вещества, и покидают его молекулы РНК, отработанные белки, субъ единицы рибосом. Таким образом, функциями ядерной оболочки являются отделение содержимого ядра от цитоплазмы, а также регуляция обмена веществ между ядром и цитоплазмой.
Нуклеоплазмой называют содержимое ядра, в которое погружены хроматин и ядрышко. Она представляет собой коллоидный раствор, по химическому составу напоминающий цитоплазму. Ферменты нуклеоплазмы катализируют обмен аминокислот, нуклеотидов, белков и др. Нуклеоплазма связана с гиалоплазмой через ядерные поры. Функции нуклеоплазмы, как и гиалоплазмы, состоят в обеспечении взаимосвязи всех структурных компонентов ядра и осуществлении ряда ферментных реакций.
Хроматином называют совокупность тонких нитей и гранул, погруженных в нуклеоплазму. Выявить его можно только при окрашивании, так как коэффициенты преломления хроматина и нуклеоплазмы приблизительно одинаковы. Нитчатый компонент хроматина называют эухроматином, а гранулярный — гетерохроматином. Эухроматин слабо уплотнен, поскольку с него считывается наследственная информация, тогда как более спирализованный гетерохроматин является генетически неактивным.
Хроматин представляет собой структурное видоизменение хромосом в неделящемся ядре. Таким образом, хромосомы постоянно присутствуют в ядре, изменяется лишь их состояние в зависимости от функции, которую ядро выполняет в данный момент.
В состав хроматина в основном входят белки-нуклеопротеины (дезоксирибонуклеопротеины и рибонуклеопротеины), а также ферменты, важнейшие из которых связаны с синтезом нуклеиновых кислот, и некоторые другие вещества.
Функции хроматина состоят, во-первых, в синтезе специфических для данного организма нуклеиновых кислот, которые направляют синтез специфических белков, во-вторых, в передаче наследственных свойств от материнской клетки дочерним, для чего хроматиновые нити в процессе деления упаковываются в хромосомы.
Ядрышко — сферическое, хорошо заметное под микроскопом тельце диаметром 1–3 мкм. Оно формируется на участках хроматина, в которых закодирована информация о структуре рРНК и белках рибосом. Ядрышко в ядре часто одно, однако в тех клетках, где происходят интенсивные процессы жизнедеятельности, ядрышек может быть два и более. Функции ядрышек — синтез рРНК и сборка субъединиц рибосом путем объединения рРНК с белками, поступающими из цитоплазмы.
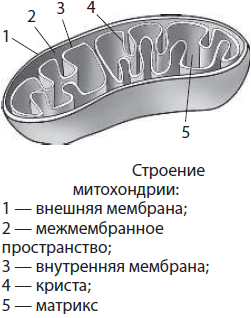
Митохондрии — двумембранные органоиды округлой, овальной или палочковидной формы, хотя встречаются и спиралевидные (в сперматозоидах). Диаметр митохондрий составляет до 1 мкм, а длина — до 7 мкм. Пространство внутри митохондрий заполнено матриксом. Матрикс — это основное вещество митохондрий. В него погружены кольцевая молекула ДНК и рибосомы. Наружная мембрана митохондрий гладкая, она непроницаема для многих веществ. Внутренняя мембрана имеет выросты — кристы, увеличивающие площадь поверхности мембран для протекания химических реакций. На поверхности мембраны расположены многочисленные белковые комплексы, составляющие так называемую дыхательную цепь, а также грибовидные ферменты АТФ-синтетазы. В митохондриях протекает аэробный этап дыхания, в ходе которого происходит синтез АТФ.
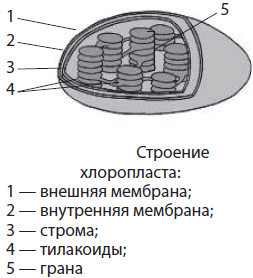
Пластиды — крупные двумембранные органоиды, характерные только для растительных клеток. Внутреннее пространство пластид заполнено стромой, или матриксом. В строме находится более или менее развитая система мембранных пузырьков — тилакоидов, которые собраны в стопки — граны, а также собственная кольцевая молекула ДНК и рибосомы. Различают четыре основных типа пластид: хлоропласты, хромопласты, лейкопласты и пропластиды.
Хлоропласты — это зеленые пластиды диаметром 3–10 мкм, хорошо различимые под микроскопом. Они содержатся только в зеленых частях растений — листьях, молодых стеблях, цветках и плодах. Хлоропласты в основном имеют овальную или эллипсоидную формы, но могут быть также чашевидными, спиралевидными и даже лопастными. Количество хлоропластов в клетке в среднем составляет от 10 до 100 штук. Однако, например, у некоторых водорослей он может быть один, иметь значительные размеры и сложную форму — тогда его называют хроматофором. В других случаях количество хлоропластов может достигать нескольких сотен, при этом их размеры невелики. Окраска хлоропластов обусловлена основным пигментом фотосинтеза — хлорофиллом, хотя в них содержатся и дополнительные пигменты — каротиноиды. Каротиноиды становятся заметными только осенью, когда хлорофилл в стареющих листьях разрушается. Основной функцией хлоропластов является фотосинтез. Световые реакции фотосинтеза протекают на мембранах тилакоидов, на которых закреплены молекулы хлорофилла, а темновые реакции — в строме, где содержатся многочисленные ферменты.
Хромопласты — это желтые, оранжевые и красные пластиды, содержащие пигменты каротиноиды. Форма хромопластов может также существенно варьировать: они бывают трубчатыми, сферическими, кристаллическими и др. Хромопласты придают окраску цветкам и плодам растений, привлекая опылителей и распространителей семян и плодов.
Лейкопласты — это белые или бесцветные пластиды в основном округлой или овальной формы. Они распространены в нефотосинтезирующих частях растений, например в кожице листа, клубнях картофеля и т. д. В них откладываются в запас питательные вещества, чаще всего крахмал, но у некоторых растений это могут быть белки или масло.
Пластиды образуются в растительных клетках из пропластид, которые имеются уже в клетках образовательной ткани и представляют собой небольшие двумембранные тельца. На ранних этапах развития разные виды пластид способны превращаться друг в друга: при попадании на свет лейкопласты клубня картофеля и хромопласты корнеплода моркови зеленеют.
Пластиды и митохондрии называют полуавтономными органоидами клетки, так как они имеют собственные молекулы ДНК и рибосомы, осуществляют синтез белка и делятся независимо от деления клеток. Эти особенности объясняются происхождением от одноклеточных прокариотических организмов. Однако «самостоятельность » митохондрий и пластид является ограниченной, так как их ДНК содержит слишком мало генов для свободного существования, остальная же информация закодирована в хромосомах ядра, что позволяет ему контролировать данные органоиды.
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭР), — это одномембранный органоид, представляющий собой сеть мембранных полостей и канальцев, занимающих до 30 % содержимого цитоплазмы. Диаметр канальцев ЭПС составляет около 25–30 нм. Различают два вида ЭПС — шероховатую и гладкую. Шероховатая ЭПС несет рибосомы, на ней происходит синтез белков. Гладкая ЭПС лишена рибосом. Ее функция — синтез липидов и углеводов, а также транспорт, запасание и обезвреживание токсических веществ. Она особенно развита в тех клетках, где происходят интенсивные процессы обмена веществ, например в клетках печени — гепатоцитах — и волокнах скелетных мышц. Вещества, синтезированные в ЭПС, транспортируются в аппарат Гольджи. В ЭПС происходит также сборка мембран клетки, однако их формирование завершается в аппарате Гольджи.
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид, образованный системой плоских цистерн, канальцев и отшнуровывающихся от них пузырьков. Структурной единицей аппарата Гольджи является диктиосома — стопка цистерн, на один полюс которой приходят вещества из ЭПС, а с противоположного полюса, подвергшись определенным превращениям, они упаковываются в пузырьки и направляются в другие части клетки. Диаметр цистерн — порядка 2 мкм, а мелких пузырьков — около 20–30 мкм. Основные функции комплекса Гольджи — синтез некоторых веществ и модификация (изменение) белков, липидов и углеводов, поступающих из ЭПС, окончательное формирование мембран, а также транспорт веществ по клетке, обновление ее структур и образование лизосом. Свое название аппарат Гольджи получил в честь итальянского ученого Камилло Гольджи, впервые обнаружившего данный органоид (1898).
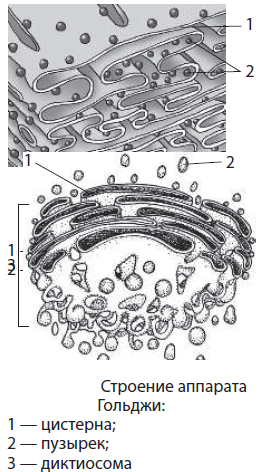
Лизосомы — небольшие одномембранные органоиды до 1 мкм в диаметре, в которых содержатся гидролитические ферменты, участвующие во внутриклеточном пищеварении. Мембраны лизосом слабопроницаемы для этих ферментов, поэтому выполнение лизосомами своих функций происходит очень точно и адресно. Так, они принимают активное участие в процессе фагоцитоза, образуя пищеварительные вакуоли, а в случае голодания или повреждения определенных частей клетки переваривают их, не затрагивая иных. Недавно была открыта роль лизосом в процессах клеточной гибели.
Вакуоль — это полость в цитоплазме растительных и животных клеток, ограниченная мембраной и заполненная жидкостью. В клетках простейших обнаруживаются пищеварительные и сократительные вакуоли. Первые принимают участие в процессе фагоцитоза, так как в них происходит расщепление питательных веществ. Вторые обеспечивают поддержание водно-солевого баланса за счет осморегуляции. У многоклеточных животных в основном встречаются пищеварительные вакуоли.
В растительных клетках вакуоли присутствуют всегда, они окружены специальной мембраной и заполнены клеточным соком. Мембрана, окружающая вакуоль, по химическому составу, строению и выполняемым функциям близка к плазматической мембране. Клеточный сок представляет собой водный раствор различных неорганических и органических веществ, в том числе минеральных солей, органических кислот, углеводов, белков, гликозидов, алкалоидов и др. Вакуоль может занимать до 90 % объема клетки и оттеснять ядро на периферию. Эта часть клетки выполняет запасающую, выделительную, осмотическую, защитную, лизосомную и другие функции, поскольку в ней накапливаются питательные вещества и отходы жизнедеятельности, она обеспечивает поступление воды и поддержание формы и объема клетки, а также содержит ферменты расщепления многих компонентов клетки. К тому же биологически активные вещества вакуолей способны препятствовать поеданию этих растений многими животными. У ряда растений за счет разбухания вакуолей происходит рост клетки растяжением.
Вакуоли имеются также и в клетках некоторых грибов и бактерий, однако у грибов они выполняют только функцию осморегуляции, а у цианобактерий поддерживают плавучесть и участвуют в процессах усвоения азота из воздуха.
Рибосомы — небольшие немембранные органоиды диаметром 15–20 мкм, состоящие из двух субъединиц — большой и малой. Субъединицы рибосом эукариот собираются в ядрышке, а затем транспортируются в цитоплазму. Рибосомы прокариот, митохондрий и пластид меньше по величине, чем рибосомы эукариот. В состав субъединиц рибосом входят рРНК и белки.
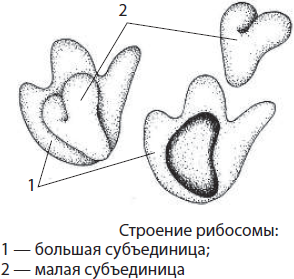
Количество рибосом в клетке может достигать нескольких десятков миллионов: в цитоплазме, митохондриях и пластидах они находятся в свободном состоянии, а на шероховатой ЭПС — в связанном. Они принимают участие в синтезе белка, в частности, осуществляют процесс трансляции — биосинтеза полипептидной цепи на молекуле иРНК. На свободных рибосомах синтезируются белки гиалоплазмы, митохондрий, пластид и собственные белки рибосом, тогда как на прикрепленных к шероховатой ЭПС рибосомах осуществляется трансляция белков для выведения из клеток, сборки мембран, образования лизосом и вакуолей.
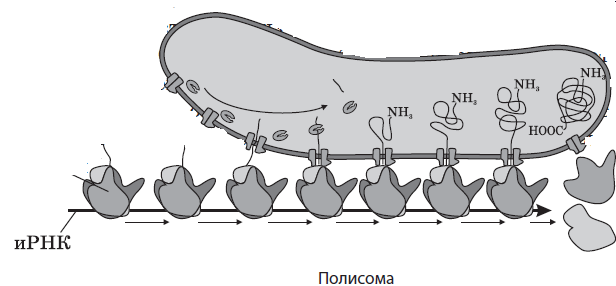
Рибосомы могут находиться в гиалоплазме поодиночке или собираться в группы при одновременном синтезе на одной иРНК сразу нескольких полипептидных цепей. Такие группы рибосом называются полирибосомами, или полисомами.
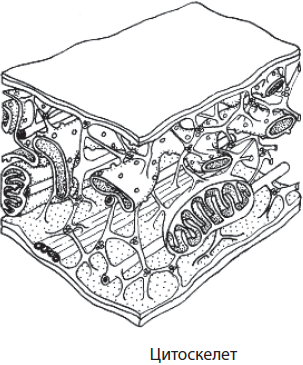
Микротрубочки — это цилиндрические полые немембранные органоиды, которые пронизывают всю цитоплазму клетки. Их диаметр составляет около 25 нм, толщина стенки — 6–8 нм. Они образованы многочисленными молекулами белка тубулина, которые сначала формируют 13 нитей, напоминающих бусы, а затем собираются в микротрубочку. Микротрубочки образуют цитоплазматическую сеть, которая придает клетке форму и объем, связывают плазматическую мембрану с другими частями клетки, обеспечивают транспорт веществ по клетке, принимают участие в движении клетки и внутриклеточных компонентов, а также в делении генетического материала. Они входят в состав клеточного центра и органоидов движения — жгутиков и ресничек.
Микрофиламенты, или микронити, также являются немембранными органоидами, однако они имеют нитевидную форму и образованы не тубулином, а актином. Они принимают участие в процессах мембранного транспорта, межклеточном узнавании, делении цитоплазмы клетки и в ее движении. В мышечных клетках взаимодействие актиновых микрофиламентов с миозиновыми нитями обеспечивает сокращение.
Микротрубочки и микрофиламенты образуют внутренний скелет клетки — цитоскелет. Он представляет собой сложную сеть волокон, обеспечивающих механическую опору для плазматической мембраны, определяет форму клетки, расположение клеточных органоидов и их перемещение в процессе деления клетки.
Клеточный центр — немембранный органоид, располагающийся в животных клетках вблизи ядра; в растительных клетках он отсутствует. Его длина составляет около 0.2–0.3 мкм, а диаметр — 0.1–0.15 мкм. Клеточный центр образован двумя центриолями, лежащими во взаимно перпендикулярных плоскостях, и лучистой сферой из микротрубочек. Каждая центриоль образована девятью группами микротрубочек, собранных по три, т. е. триплетами. Клеточный центр принимает участие в процессах сборки микротрубочек, делении наследственного материала клетки, а также в образовании жгутиков и ресничек.
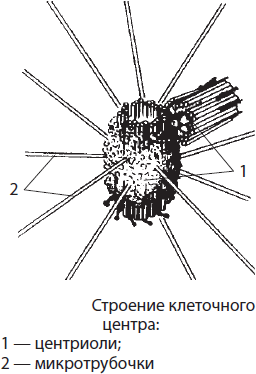
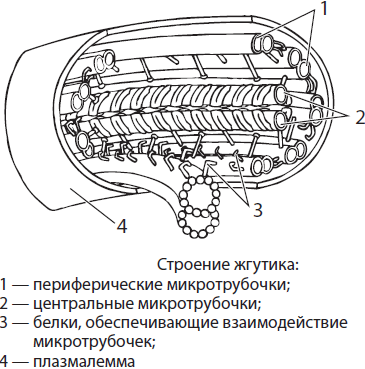
Органоиды движения. Жгутики и реснички представляют собой выросты клетки, покрытые плазмалеммой. Основу этих органоидов составляют девять пар микротрубочек, расположенных по периферии, и две свободные микротрубочки в центре. Микротрубочки связаны между собой различными белками, обеспечивающими их согласованное отклонение от оси — колебание. Колебания энергозависимы, то есть на этот процесс тратится энергия макроэргических связей АТФ. Восстановление утраченных жгутиков и ресничек является функцией базальных телец, или кинетосом, расположенных в их основании.
Длина ресничек составляет около 10–15 нм, а жгутиков — 20–50 мкм. За счет строго направленных движений жгутиков и ресничек осуществляется не только движение одноклеточных животных, сперматозоидов и др., но и происходит очистка дыхательных путей, продвижение яйцеклетки по маточным трубам, поскольку все эти части организма человека выстланы реснитчатым эпителием.
Включения
Включения — это непостоянные компоненты клетки, которые образуются и исчезают в процессе ее жизнедеятельности. К ним относят как запасные вещества, например, зерна крахмала или белка в растительных клетках, гранулы гликогена в клетках животных и грибов, волютина у бактерий, капли жира во всех типах клеток, так и отходы жизнедеятельности, в частности, непереваренные в результате фагоцитоза остатки пищи, образующие так называемые остаточные тельца.
Взаимосвязь строения и функций частей и органоидов клетки — основа ее целостности
Каждая из частей клетки, с одной стороны, является обособленной структурой со специфическим строением и функциями, а с другой — компонентом более сложной системы, называемой клеткой. Бульшая часть наследственной информации эукариотической клетки сосредоточена в ядре, однако само ядро не в состоянии обеспечить ее реализацию, поскольку для этого необходимы как минимум цитоплазма, выступающая как основное вещество, и рибосомы, на которых и происходит этот синтез. Большинство рибосом расположено на гранулярной эндоплазматической сети, откуда белки чаще всего транспортируются в комплекс Гольджи, а затем после модификации — в те части клетки, для которых они предназначены, или выводятся наружу. Мембранные упаковки белков и углеводов могут встраиваться в мембраны органоидов и цитоплазматическую мембрану, обеспечивая их постоянное обновление. От комплекса Гольджи отшнуровываются также выполняющие важнейшие функции лизосомы и вакуоли. Например, без лизосом клетки быстро превратились бы в свое образную свалку отработанных молекул и структур.
Протекание всех этих процессов требует энергии, вырабатываемой митохондриями, а у растений — и хлоропластами. И хотя эти органоиды являются относительно автономными, т. к. имеют собственные молекулы ДНК, часть их белков все равно кодируется ядерным геномом и синтезируется в цитоплазме.
Таким образом, клетка представляет собой неразрывное единство составляющих ее компонентов, каждый из которых выполняет свою уникальную функцию.
Обмен веществ и превращения энергии — свойства живых организмов. Энергетический и пластический обмен, их взаимосвязь. Стадии энергетического обмена. Брожение и дыхание. Фотосинтез, его значение, космическая роль. Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь. Хемосинтез. Роль хемосинтезирующих бактерий на Земле
Обмен веществ и превращения энергии — свойства живых организмов
Клетку можно уподобить миниатюрной химической фабрике, на которой происходят сотни и тысячи химических реакций.
Обмен веществ — совокупность химических превращений, направленных на сохранение и самовоспроизведение биологических систем.
Он включает в себя поступление веществ в организм в процессе питания и дыхания, внутриклеточный обмен веществ, или метаболизм, а также выделение конечных продуктов обмена.
Обмен веществ неразрывно связан с процессами превращения одних видов энергии в другие. Например, в процессе фотосинтеза световая энергия запасается в виде энергии химических связей сложных органических молекул, а в процессе дыхания она высвобождается и расходуется на синтез новых молекул, механическую и осмотическую работу, рассеивается в виде тепла и т. д.
Протекание химических реакций в живых организмах обеспечивается благодаря биологическим катализаторам белковой природы — ферментам, или энзимам. Как и другие катализаторы, ферменты ускоряют протекание химических реакций в клетке в десятки и сотни тысяч раз, а иногда и вообще делают их возможными, но не изменяют при этом ни природы, ни свойств конечного продукта (продуктов) реакции и не изменяются сами. Ферменты могут быть как простыми, так и сложными белками, в состав которых, кроме белковой части, входит и небелковая — кофактор (кофермент). Примерами ферментов являются амилаза слюны, расщепляющая полисахариды при длительном пережевывании, и пепсин, обеспечивающий переваривание белков в желудке.
Ферменты отличаются от катализаторов небелковой природы высокой специфичностью действия, значительным увеличением с их помощью скорости реакции, а также возможностью регуляции действия за счет изменения условий протекания реакции либо взаимодействия с ними различных веществ. К тому же и условия, в которых протекает ферментный катализ, существенно отличаются от тех, при которых идет неферментный: оптимальной для функционирования ферментов в организме человека является температура $37°С$, давление должно быть близким к атмосферному, а $рН$ среды может существенно колебаться. Так, для амилазы необходима щелочная среда, а для пепсина — кислая.
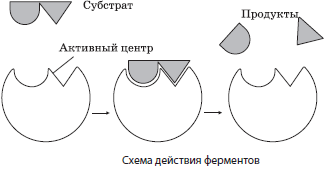
Механизм действия ферментов заключается в снижении энергии активации веществ (субстратов), вступающих в реакцию, за счет образования промежуточных фермент-субстратных комплексов.
Энергетический и пластический обмен, их взаимосвязь
Метаболизм складывается из двух одновременно протекающих в клетке процессов: пластического и энергетического обменов.
Пластический обмен (анаболизм, ассимиляция) представляет собой совокупность реакций синтеза, которые идут с затратой энергии АТФ. В процессе пластического обмена синтезируются органические вещества, необходимые клетке. Примером реакций пластического обмена являются фотосинтез, биосинтез белка и репликация (самоудвоение) ДНК.
Энергетический обмен (катаболизм, диссимиляция) — это совокупность реакций расщепления сложных веществ до более простых. В результате энергетического обмена выделяется энергия, запасаемая в виде АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.
Пластический и энергетический обмены неразрывно связаны, поскольку в процессе пластического обмена синтезируются органические вещества и для этого необходима энергия АТФ, а в процессе энергетического обмена органические вещества расщепляются и высвобождается энергия, которая затем будет израсходована на процессы синтеза.
Энергию организмы получают в процессе питания, а высвобождают ее и переводят в доступную форму в основном в процессе дыхания. По способу питания все организмы делятся на автотрофов и гетеротрофов. Автотрофы способны самостоятельно синтезировать органические вещества из неорганических, а гетеротрофы используют исключительно готовые органические вещества.
Стадии энергетического обмена
Несмотря на всю сложность реакций энергетического обмена, его условно подразделяют на три этапа: подготовительный, анаэробный (бескислородный) и аэробный (кислородный).
На подготовительном этапе молекулы полисахаридов, липидов, белков, нуклеиновых кислот распадаются на более простые, например, глюкозу, глицерин и жирные кислоты, аминокислоты, нуклеотиды и др. Этот этап может протекать непосредственно в клетках либо в кишечнике, откуда расщепленные вещества доставляются с током крови.
Анаэробный этап энергетического обмена сопровождается дальнейшим расщеплением мономеров органических соединений до еще более простых промежуточных продуктов, например, пировиноградной кислоты, или пирувата. Он не требует присутствия кислорода, и для многих организмов, обитающих в иле болот или в кишечнике человека, является единственным способом получения энергии. Анаэробный этап энергетического обмена протекает в цитоплазме.
Бескислородному расщеплению могут подвергаться различные вещества, однако довольно часто субстратом реакций оказывается глюкоза. Процесс ее бескислородного расщепления называется гликолизом. При гликолизе молекула глюкозы теряет четыре атома водорода, т. е. окисляется, при этом образуются две молекулы пировиноградной кислоты, две молекулы АТФ и две молекулы восстановленного переносчика водорода $НАДН + Н^{+}$:
$С_6Н_{12}О_6 + 2Н_3РО_4 + 2АДФ + 2НАД → 2С_3Н_4О_3 + 2АТФ + 2НАДН + Н^{+} + 2Н_2О$.
Образование АТФ из АДФ происходит вследствие прямого переноса фосфат-аниона с предварительно фосфорилированного сахара и называется субстратным фосфорилированием.
Аэробный этап энергетического обмена может происходить только в присутствии кислорода, при этом промежуточные соединения, образовавшиеся в процессе бескислородного расщепления, окисляются до конечных продуктов (углекислого газа и воды) и выделяется большая часть энергии, запасенной в химических связях органических соединений. Она переходит в энергию макроэргических связей 36 молекул АТФ. Этот этап также называется тканевым дыханием. В случае отсутствия кислорода промежуточные соединения превращаются в другие органические вещества, и этот процесс называется брожением.
Дыхание
Механизм клеточного дыхания схематически изображен на рис.
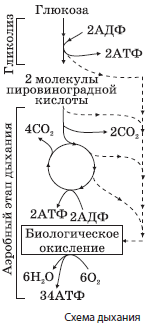
Аэробное дыхание происходит в митохондриях, при этом пировиноградная кислота сначала утрачивает один атом углерода, что сопровождается синтезом одного восстановительного эквивалента $НАДН + Н^{+}$ и молекулы ацетилкофермента А (ацетил-КоА):
$С_3Н_4О_3 + НАД + Н~КоА → СН_3СО~КоА + НАДН + Н^{+} + СО_2↑$.
Ацетил-КоА в матриксе митохондрий вовлекается в цепь химических реакций, совокупность которых называется циклом Кребса (циклом трикарбоновых кислот, циклом лимонной кислоты). В ходе этих превращений образуется две молекулы АТФ, ацетил-КоА полностью окисляется до углекислого газа, а его ионы водорода и электроны присоединяются к переносчикам водорода $НАДН + Н^{+}$ и $ФАДН_2$. Переносчики транспортируют протоны водорода и электроны к внутренним мембранам митохондрий, образующим кристы. При помощи белков-переносчиков протоны водорода нагнетаются в межмембранное пространство, а электроны передаются по так называемой дыхательной цепи ферментов, расположенной на внутренней мембране митохондрий, и сбрасываются на атомы кислорода:
$O_2+2e^{-}→O_2^-$.
Следует отметить, что некоторые белки дыхательной цепи содержат железо и серу.
Из межмембранного пространства протоны водорода транспортируются обратно в матрикс митохондрий с помощью специальных ферментов — АТФ-синтаз, а выделяющаяся при этом энергия расходуется на синтез 34 молекул АТФ из каждой молекулы глюкозы. Этот процесс называется окислительным фосфорилированием. В матриксе митохондрий протоны водорода реагируют с радикалами кислорода с образованием воды:
$4H^{+}+O_2^-→2H_2O$.
Совокупность реакций кислородного дыхания может быть выражена следующим образом:
$2С_3Н_4О_3 + 6О_2 + 36Н_3РО_4 + 36АДФ → 6СО_2↑ + 38Н_2О + 36АТФ.$
Суммарное уравнение дыхания выглядит таким образом:
$С_6Н_{12}О_6 + 6О_2 + 38Н_3РО_4 + 38АДФ → 6СО_2↑ + 40Н_2О + 38АТФ.$
Брожение
В отсутствие кислорода или при его недостатке происходит брожение. Брожение является эволюционно более ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку в результате брожения образуются органические вещества, все еще богатые энергией. Различают несколько основных видов брожения: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода в ходе брожения пировиноградная кислота восстанавливается до молочной кислоты, при этом образовавшиеся ранее восстановительные эквиваленты расходуются, и остаются всего две молекулы АТФ:
$2С_3Н_4О_3 + 2НАДН + Н^{+} → 2С_3Н_6О_3 + 2НАД$.
При брожении с помощью дрожжевых грибов пировиноградная кислота в присутствии кислорода превращается в этиловый спирт и оксид углерода (IV):
$С_3Н_4О_3 + НАДН + Н^{+} → С_2Н_5ОН + СО_2↑ + НАД^{+}$.
При брожении с помощью микроорганизмов из пировиноградной кислоты могут образоваться также уксусная, масляная, муравьиная кислоты и др.
АТФ, полученная в результате энергетического обмена, расходуется в клетке на различные виды работы: химическую, осмотическую, электрическую, механическую и регуляторную. Химическая работа заключается в биосинтезе белков, липидов, углеводов, нуклеиновых кислот и других жизненно важных соединений. К осмотической работе относят процессы поглощения клеткой и выведения из нее веществ, которые во внеклеточном пространстве находятся в концентрациях, больших, чем в самой клетке. Электрическая работа тесно взаимосвязана с осмотической, поскольку именно в результате перемещения заряженных частиц через мембраны формируется заряд мембраны и приобретаются свойства возбудимости и проводимости. Механическая работа сопряжена с движением веществ и структур внутри клетки, а также клетки в целом. К регуляторной работе относят все процессы, направленные на координацию процессов в клетке.
Фотосинтез, его значение, космическая роль
Фотосинтезом называют процесс преобразования энергии света в энергию химических связей органических соединений с участием хлорофилла.
В результате фотосинтеза образуется около 150 млрд тонн органического вещества и приблизительно 200 млрд тонн кислорода ежегодно. Этот процесс обеспечивает круговорот углерода в биосфере, не давая накапливаться углекислому газу и препятствуя тем самым возникновению парникового эффекта и перегреву Земли. Образующиеся в результате фотосинтеза органические вещества не расходуются другими организмами полностью, значительная их часть в течение миллионов лет образовала залежи полезных ископаемых (каменного и бурого угля, нефти). В последнее время в качестве топлива начали использовать также рапсовое масло («биодизель») и спирт, полученный из растительных остатков. Из кислорода под действием электрических разрядов образуется озон, который формирует озоновый экран, защищающий все живое на Земле от губительного действия ультрафиолетовых лучей.
Наш соотечественник, выдающийся физиолог растений К. А. Тимирязев (1843–1920) назвал роль фотосинтеза «космической», поскольку он связывает Землю с Солнцем (космосом), обеспечивая приток энергии на планету.
Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь
В 1905 году английский физиолог растений Ф. Блэкмен обнаружил, что скорость фотосинтеза не может увеличиваться беспредельно, какой-то фактор ограничивает ее. На основании этого он выдвинул предположение о наличии двух фаз фотосинтеза: световой и темновой. При низкой интенсивности освещения скорость световых реакций возрастает пропорционально нарастанию силы света, и, кроме того, данные реакции не зависят от температуры, поскольку для их протекания не нужны ферменты. Световые реакции протекают на мембранах тилакоид.
Скорость темновых реакций, напротив, возрастает с повышением температуры, однако по достижении температурного порога в $30°С$ этот рост прекращается, что свидетельствует о ферментативном характере указанных превращений, происходящих в строме. Следует отметить, что свет также оказывает на темновые реакции определенное влияние, несмотря на то, что они называются темновыми.
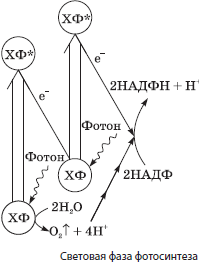
Световая фаза фотосинтеза протекает на мембранах тилакоидов, несущих несколько типов белковых комплексов, основными из которых являются фотосистемы I и II, а также АТФсинтаза. В состав фотосистем входят пигментные комплексы, в которых, кроме хлорофилла, присутствуют и каротиноиды. Каротиноиды улавливают свет в тех областях спектра, в которых этого не делает хлорофилл, а также защищают хлорофилл от разрушения светом высокой интенсивности.
Кроме пигментных комплексов, фотосистемы включают и ряд белков-акцепторов электронов, которые последовательно передают друг другу электроны от молекул хлорофилла. Последовательность этих белков называется электронтранспортной цепью хлоропластов.
С фотосистемой II также ассоциирован специальный комплекс белков, который обеспечивает выделение кислорода в процессе фотосинтеза. Этот кислородвыделяющий комплекс содержит ионы марганца и хлора.
В световой фазе кванты света, или фотоны, попадающие на молекулы хлорофилла, расположенные на мембранах тилакоидов, переводят их в возбужденное состояние, характеризующееся более высокой энергией электронов. При этом возбужденные электроны от хлорофилла фотосистемы I передаются через цепь посредников на переносчик водорода НАДФ, присоединяющий при этом протоны водорода, всегда имеющиеся в водном растворе:
$НАДФ + 2e^{-} + 2H^{+} → НАДФН + Н^{+}$.
Восстановленный $НАДФН + Н^{+}$ будет впоследствии использован в темновой стадии. Электроны от хлорофилла фотосистемы II также передаются по электронтранспортной цепи, однако они заполняют «электронные дырки» хлорофилла фотосистемы I. Недостаток электронов в хлорофилле фотосистемы II заполняется за счет отнимания у молекул воды, которое происходит с участием уже упоминавшегося выше кислородвыделяющего комплекса. В результате разложения молекул воды, которое называется фотолизом, образуются протоны водорода и выделяется молекулярный кислород, являющийся побочным продуктом фотосинтеза:
$H_2O → 2H^{+} + 2e^{-} + {1}/{2}O_2↑$.
Генетическая информация в клетке. Гены, генетический код и его свойства. Матричный характер реакций биосинтеза. Биосинтез белка и нуклеиновых кислот
Генетическая информация в клетке
Воспроизведение себе подобных является одним из фундаментальных свойств живого. Благодаря этому явлению существует сходство не только между организмами, но и между отдельными клетками, а также их органоидами (митохондриями и пластидами). Материальной основой этого сходства является передача зашифрованной в последовательности нуклеотидов ДНК генетической информации, которая осуществляется благодаря процессам репликации (самоудвоения) ДНК. Реа лизуются все признаки и свойства клеток и организмов благодаря белкам, структуру которых в первую очередь и определяют последовательности нуклеотидов ДНК. Поэтому первостепенное значение в процессах метаболизма играет именно биосинтез нуклеиновых кислот и белка. Структурной единицей наследственной информации является ген.
Гены, генетический код и его свойства
Наследственная информация в клетке не является монолитной, она разбита на отдельные «слова» — гены.
Ген — это элементарная единица генетической информации.
Работы по программе «Геном человека», которые проводились одновременно в нескольких странах и были завершены в начале нынешнего века, дали нам понимание того, что у человека всего около 25–30 тыс. генов, но информация с большей части нашей ДНК не считывается никогда, так как в ней содержится огромное количество бессмысленных участков, повторов и генов, кодирующих признаки, утратившие значение для человека (хвост, оволосение тела и др.). Кроме того, был расшифрован ряд генов, отвечающих за развитие наследственных заболеваний, а также генов-мишеней лекарственных препаратов. Однако практическое применение результатов, полученных в ходе реализации данной программы, откладывается до тех пор, пока не будут расшифрованы геномы большего количества людей и станет понятно, чем же все-таки они различаются.
Гены, кодирующие первичную структуру белка, рибосомальной или транспортной РНК называются структурными, а гены, обеспечивающие активацию или подавление считывания информации со структурных генов, — регуляторными. Однако даже структурные гены содержат регуляторные участки.
Наследственная информация организмов зашифрована в ДНК в виде определенных сочетаний нуклеотидов и их последовательности — генетического кода. Его свойствами являются: триплетность, специфичность, универсальность, избыточность и неперекрываемость. Кроме того, в генетическом коде отсутствуют знаки препинания.
Каждая аминокислота закодирована в ДНК тремя нуклеотидами — триплетом, например, метионин закодирован триплетом ТАЦ, то есть код триплетен. С другой стороны, каждый триплет кодирует только одну аминокислоту, в чем заключается его специфичность или однозначность. Генетический код универсален для всех живых организмов, то есть наследственная информация о белках человека может считываться бактериями и наоборот. Это свидетельствует о единстве происхождения органического мира. Однако 64 комбинациям нуклеотидов по три соответствует только 20 аминокислот, вследствие чего одну аминокислоту может кодировать 2–6 триплетов, то есть генетический код избыточен, или вырожден. Три триплета не имеют соответствующих аминокислот, их называют стоп-кодонами, так как они обозначают окончание синтеза полипептидной цепи.
Последовательность оснований в триплетах ДНК и кодируемые ими аминокислоты
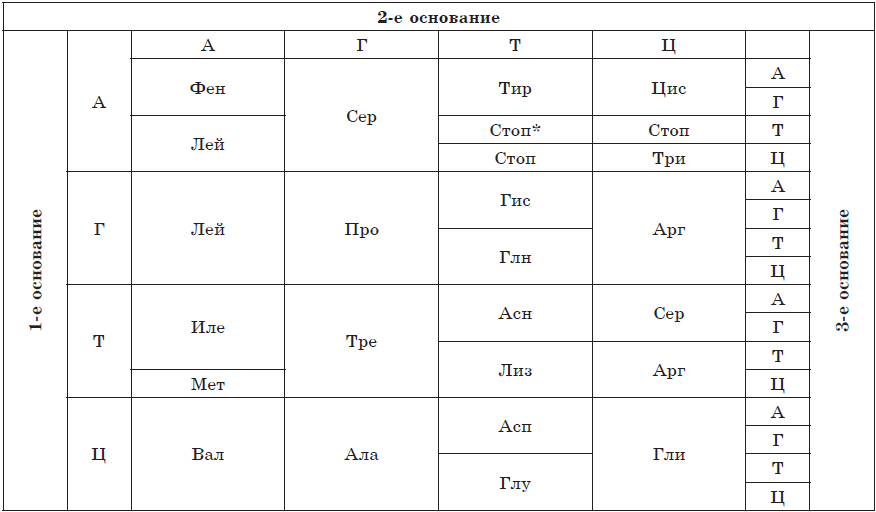
*Стоп-кодон, означающий конец синтеза полипептидной цепи.
Сокращения названий аминокислот:
Ала — аланин
Арг — аргинин
Асн — аспарагин
Асп — аспарагиновая кислота
Вал — валин
Гис — гистидин
Гли — глицин
Глн — глутамин
Глу — глутаминовая кислота
Иле — изолейцин
Лей — лейцин
Лиз — лизин
Мет — метионин
Про — пролин
Сер — серин
Тир — тирозин
Тре — треонин
Три — триптофан
Фен — фенилаланин
Цис — цистеин
Если начать считывание генетической информации не с первого нуклеотида в триплете, а со второго, то произойдет не только сдвижка рамки считывания — синтезированный таким образом белок будет совсем иным не только по последовательности нуклеотидов, но и по структуре и свойствам. Между триплетами отсутствуют какие бы то ни было знаки препинания, поэтому нет никаких препятствий для сдвижки рамки считывания, что открывает простор для возникновения и сохранения мутаций.
Матричный характер реакций биосинтеза
Клетки бактерий способны удваиваться каждые 20–30 минут, а клетки эукариот — каждые сутки и даже чаще, что требует высокой скорости и точности репликации ДНК. Кроме того, каждая клетка содержит сотни и тысячи копий многих белков, особенно ферментов, следовательно, для их воспроизведения неприемлем «штучный» способ их производства. Более прогрессивным способом является штамповка, которая позволяет получить многочисленные точные копии продукта и к тому же снизить его себестоимость. Для штамповки необходима матрица, с которой осуществляется оттиск.
В клетках принцип матричного синтеза заключается в том, что новые молекулы белков и нуклеиновых кислот синтезируются в соответствии с программой, заложенной в структуре ранее существовавших молекул тех же нуклеиновых кислот (ДНК или РНК).
Биосинтез белка и нуклеиновых кислот
Репликация ДНК. ДНК представляет собой двухцепочечный биополимер, мономерами которого являются нуклеотиды. Если бы биосинтез ДНК происходил по принципу ксерокопирования, то неизбежно возникали бы многочисленные искажения и погрешности в наследственной информации, которые в конечном итоге привели бы к гибели новых организмов. Поэтому процесс удвоения ДНК происходит иным, полуконсервативным способом: молекула ДНК расплетается, и на каждой из цепей синтезируется новая цепь по принципу комплементарности. Процесс самовоспроизведения молекулы ДНК, обеспечивающий точное копирование наследственной информации и передачу ее из поколения в поколение, называется репликацией (от лат. репликацио — повторение). В результате репликации образуются две абсолютно точные копии материнской молекулы ДНК, каждая из которых несет по одной копии материнской.
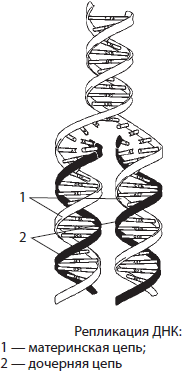
Процесс репликации на самом деле крайне сложен, так как в нем участвует целый ряд белков. Одни из них раскручивают двойную спираль ДНК, другие разрывают водородные связи между нуклеотидами комплементарных цепей, третьи (например, фермент ДНК-полимераза) подбирают по принципу комплементарности новые нуклеотиды и т. д. Образовавшиеся в результате репликации две молекулы ДНК в процессе деления расходятся по двум вновь образующимся дочерним клеткам.
Ошибки в процессе репликации возникают крайне редко, однако если они и происходят, то очень быстро устраняются как ДНК-полимеразами, так и специальными ферментами репарации, поскольку любая ошибка в последовательности нуклеотидов может привести к необратимому изменению структуры и функций белка и, в конечном итоге, неблагоприятно сказаться на жизнеспособности новой клетки или даже особи.
Биосинтез белка. Как образно выразился выдающийся философ XIX века Ф. Энгельс: «Жизнь есть форма существования белковых тел». Структура и свойства белковых молекул определяются их первичной структурой, т. е. последовательностью аминокислот, зашифрованной в ДНК. От точности воспроизведения этой информации зависит не только существование самого полипептида, но и функционирование клетки в целом, поэтому процесс синтеза белка имеет огромное значение. Он, по-видимому, является самым сложным процессом синтеза в клетке, поскольку здесь участвует до трехсот различных ферментов и других макромолекул. Кроме того, он протекает с высокой скоростью, что требует еще большей точности.
В биосинтезе белка выделяют два основных этапа: транскрипцию и трансляцию.
Транскрипция (от лат. транскрипцио — переписывание) — это биосинтез молекул иРНК на матрице ДНК.
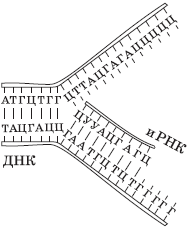
Поскольку молекула ДНК содержит две антипараллельных цепи, то считывание информации с обеих цепей привело бы к образованию совершенно различных иРНК, поэтому их биосинтез возможен только на одной из цепей, которую называют кодирующей, или кодогенной, в отличие от второй, некодирующей, или некодогенной. Обеспечивает процесс переписывания специальный фермент РНК-полимераза, который подбирает нуклеотиды РНК по принципу комплементарности. Этот процесс может протекать как в ядре, так и в органоидах, имеющих собственную ДНК, — митохондриях и пластидах.
Синтезированные в процессе транскрипции молекулы иРНК проходят сложный процесс подготовки к трансляции (митохондриальные и пластидные иРНК могут оставаться внутри органоидов, где и происходит второй этап биосинтеза белка). В процессе созревания иРНК к ней присоединяются первые три нуклеотида (АУГ) и хвост из адениловых нуклеотидов, длина которого определяет, сколько копий белка может синтезироваться на данной молекуле. Только потом зрелые иРНК покидают ядро через ядерные поры.
Параллельно в цитоплазме происходит процесс активации аминокислот, в ходе которого аминокислота присоединяется к соответствующей свободной тРНК. Этот процесс катализируется специальным ферментом, на него затрачивается АТФ.
Трансляция (от лат. трансляцио — передача) — это биосинтез полипептидной цепи на матрице иРНК, при котором происходит перевод генетической информации в последовательность аминокислот полипептидной цепи.
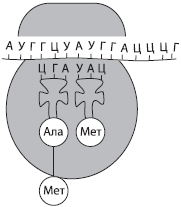
Второй этап синтеза белка чаще всего происходит в цитоплазме, например на шероховатой ЭПС. Для его протекания необходимы наличие рибосом, активация тРНК, в ходе которой они присоединяют соответствующие аминокислоты, присутствие ионов Mg2+, а также оптимальные условия среды (температура, рН, давление и т. д.).
Для начала трансляции (инициации) к готовой к синтезу молекуле иРНК присоединяется малая субъединица рибосомы, а затем по принципу комплементарности к первому кодону (АУГ) подбирается тРНК, несущая аминокислоту метионин. Лишь после этого присоединяется большая субъединица рибосомы. В пределах собранной рибосомы оказываются два кодона иРНК, первый из которых уже занят. К соседнему с ним кодону присоединяется вторая тРНК, также несущая аминокислоту, после чего между остатками аминокислот с помощью ферментов образуется пептидная связь. Рибосома передвигается на один кодон иРНК; первая из тРНК, освободившаяся от аминокислоты, возвращается в цитоплазму за следующей аминокислотой, а фрагмент будущей полипептидной цепи как бы повисает на оставшейся тРНК. К новому кодону, оказавшемуся в пределах рибосомы, присоединяется следующая тРНК, процесс повторяется и шаг за шагом полипептидная цепь удлиняется, т. е. происходит ее элонгация.
Окончание синтеза белка (терминация) происходит, как только в молекуле иРНК встретится специфическая последовательность нуклеотидов, которая не кодирует аминокислоту (стоп-кодон). После этого рибосома, иРНК и полипептидная цепь разделяются, а вновь синтезированный белок приобретает соответствующую структуру и транспортируется в ту часть клетки, где он будет выполнять свои функции.
Трансляция является весьма энергоемким процессом, поскольку на присоединение одной аминокислоты к тРНК расходуется энергия одной молекулы АТФ, еще несколько используются для продвижения рибосомы по молекуле иРНК.
Для ускорения синтеза определенных белковых молекул к молекуле иРНК могут присоединяться последовательно несколько рибосом, которые образуют единую структуру — полисому.
Клетка — генетическая единица живого. Хромосомы, их строение (форма и размеры) и функции. Число хромосом и их видовое постоянство. Соматические и половые клетки. Жизненный цикл клетки: интерфаза и митоз. Митоз — деление соматических клеток. Мейоз. Фазы митоза и мейоза. Развитие половых клеток у растений и животных. Деление клетки — основа роста, развития и размножения организмов. Роль мейоза и митоза
Клетка — генетическая единица живого
Несмотря на то, что нуклеиновые кислоты являются носителем генетической информации, реализация этой информации невозможна вне клетки, что легко доказывается на примере вирусов. Данные организмы, содержащие зачастую только ДНК или РНК, не могут самостоятельно воспроизводиться, для этого они должны использовать наследственный аппарат клетки. Даже проникнуть в клетку без помощи самой клетки они не могут, кроме как с использованием механизмов мембранного транспорта или благодаря повреждению клеток. Большинство вирусов нестабильно, они гибнут уже после нескольких часов пребывания на открытом воздухе. Следовательно, клетка является генетической единицей живого, обладающей минимальным набором компонентов для сохранения, изменения и реализации наследственной информации, а также ее передачи потомкам.
Бульшая часть генетической информации эукариотической клетки сосредоточена в ядре. Особенностью ее организации является то, что, в отличие от ДНК прокариотической клетки, молекулы ДНК эукариот не замкнуты и образуют сложные комплексы с белками — хромосомы.
Хромосомы, их строение (форма и размеры) и функции
Хромосома (от греч. хрома — цвет, окраска и сома — тело) — это структура клеточного ядра, которая содержит гены и несет определенную наследственную информацию о признаках и свойствах организма.
Иногда хромосомами называют и кольцевые молекулы ДНК прокариот. Хромосомы способны к самоудвоению, они обладают структурной и функциональной индивидуальностью и сохраняют ее в ряду поколений. Каждая клетка несет всю наследственную информацию организма, но в ней работает только небольшая часть.
Основой хромосомы является двухцепочечная молекула ДНК, упакованная с белками. У эукариот с ДНК взаимодействуют гистоновые и негистоновые белки, тогда как у прокариот гистоновые белки отсутствуют.
Лучше всего хромосомы видны под световым микроскопом в процессе деления клетки, когда они в результате уплотнения приобретают вид палочковидных телец, разделенных первичной перетяжкой — центромерой — на плечи. На хромосоме может быть также и вторичная перетяжка, которая в некоторых случаях отделяет от основной части хромосомы так называемый спутник. Концевые участки хромосом называются теломерами. Теломеры препятствуют слипанию концов хромосом и обеспечивают их прикрепление к оболочке ядра в неделящейся клетке. В начале деления хромосомы удвоены и состоят из двух дочерних хромосом — хроматид, скрепленных в центромере.
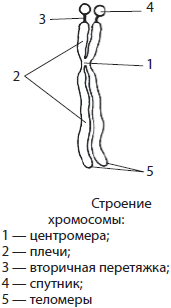
По форме различают равноплечие, неравноплечие и палочковидные хромосомы. Размеры хромосом существенно варьируют, однако средняя хромосома имеет размеры 5 $×$ 1,4 мкм.
В некоторых случаях хромосомы в результате многочисленных удвоений ДНК содержат сотни и тысячи хроматид: такие гигантские хромосомы называются политенными. Они встречаются в слюнных железах личинок дрозофилы, а также в пищеварительных железах аскариды.
Число хромосом и их видовое постоянство. Соматические и половые клетки
Согласно клеточной теории клетка является единицей строения, жизнедеятельности и развития организма. Таким образом, такие важнейшие функции живого, как рост, размножение и развитие организма обеспечиваются на клеточном уровне. Клетки многоклеточных организмов можно разделить на соматические и половые.
Соматические клетки — это все клетки тела, образующиеся в результате митотического деления.
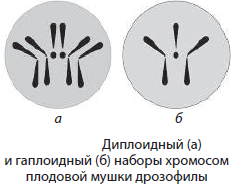
Изучение хромосом позволило установить, что для соматических клеток организма каждого биологического вида характерно постоянное число хромосом. Например, у человека их 46. Набор хромосом соматических клеток называют диплоидным (2n), или двойным.
Половые клетки, или гаметы, — это специализированные клетки, служащие для полового размножения.
В гаметах содержится всегда вдвое меньше хромосом, чем в соматических клетках (у человека — 23), поэтому набор хромосом половых клеток называется гаплоидным (n), или одинарным. Его образование связано с мейотическим делением клетки.
Количество ДНК соматических клеток обозначается как 2c, а половых — 1с. Генетическая формула соматических клеток записывается как 2n2c, а половых — 1n1с.
В ядрах некоторых соматических клеток количество хромосом может отличаться от их количества в соматических клетках. Если это различие больше на один, два, три и т. д. гаплоидных набора, то такие клетки называют полиплоидными (три-, тетра-, пентаплоидными соответственно). В таких клетках процессы метаболизма протекают, как правило, очень интенсивно.
Количество хромосом само по себе не является видоспецифическим признаком, поскольку различные организмы могут иметь равное количество хромосом, а родственные — разное. Например, у малярийного плазмодия и лошадиной аскариды по две хромосомы, а у человека и шимпанзе — 46 и 48 соответственно.
Хромосомы человека делятся на две группы: аутосомы и половые хромосомы (гетерохромосомы). Аутосом в соматических клетках человека насчитывается 22 пары, они одинаковы для мужчин и женщин, а половых хромосом только одна пара, но именно она определяет пол особи. Существует два вида половых хромосом — X и Y. Клетки тела женщины несут по две X-хромосомы, а мужчин — X и Y.
Кариотип — это совокупность признаков хромосомного набора организма (число хромосом, их форма и величина).
Условная запись кариотипа включает общее количество хромосом, половые хромосомы и возможные отклонения в наборе хромосом. Например, кариотип нормального мужчины записывается как 46, XY, а кариотип нормальной женщины — 46, XX.
Жизненный цикл клетки: интерфаза и митоз
Клетки не возникают каждый раз заново, они образуются только в результате деления материнских клеток. После разделения дочерним клеткам требуется некоторое время для формирования органоидов и приобретения соответствующей структуры, которая обеспечила бы выполнение определенной функции. Этот отрезок времени называется созреванием.
Промежуток времени от появления клетки в результате деления до ее разделения или гибели называется жизненным циклом клетки.
У эукариотических клеток жизненный цикл делится на две основные стадии: интерфазу и митоз.
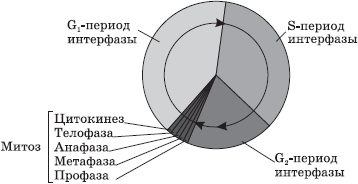
Интерфаза — это промежуток времени в жизненном цикле, в который клетка не делится и нормально функционирует. Интерфаза делится на три периода: G1-, S- и G2-периоды.
G1-период (пресинтетический, постмитотический) — это период роста и развития клетки, в который происходит активный синтез РНК, белков и других веществ, необходимых для полного жизнеобеспечения вновь образовавшейся клетки. К концу этого периода клетка может начать готовиться к удвоению ДНК.
В S-периоде (синтетическом) происходит сам процесс репликации ДНК. Единственным участком хромосомы, который не подвергается репликации, является центромера, поэтому образовавшиеся молекулы ДНК не расходятся полностью, а остаются скрепленными в ней, и в начале деления хромосома имеет X-образный вид. Генетическая формула клетки после удвоения ДНК — 2n4c. Также в S-периоде происходит удвоение центриолей клеточного центра.
G2-период (постсинтетический, премитотический) характеризуется интенсивным синтезом РНК, белков и АТФ, необходимых для процесса деления клетки, а также разделением центриолей, митохондрий и пластид. До конца интерфазы хроматин и ядрышко остаются хорошо различимыми, целостность ядерной оболочки не нарушается, а органоиды не изменяются.
Часть клеток организма способна выполнять свои функции в течение всей жизни организма (нейроны нашего головного мозга, мышечные клетки сердца), а другие существуют непродолжительное время, после чего погибают (клетки кишечного эпителия, клетки эпидермиса кожи). Следовательно, в организме должны постоянно происходить процессы деления клеток и образования новых, которые замещали бы отмершие. Клетки, способные к делению, называют стволовыми. В организме человека они находятся в красном костном мозге, в глубоких слоях эпидермиса кожи и других местах. Используя эти клетки, можно вырастить новый орган, добиться омоложения, а также клонировать организм. Перспективы использования стволовых клеток совершенно ясны, однако морально-этические аспекты этой проблемы все еще обсуждаются, поскольку в большинстве случаев используются эмбриональные стволовые клетки, полученные из убитых при аборте зародышей человека.
Продолжительность интерфазы в клетках растений и животных составляет в среднем 10– 20 часов, тогда как митоз занимает около 1–2 часов.
В ходе последовательных делений в многоклеточных организмах дочерние клетки становятся все более разнообразными, поскольку в них происходит считывание информации со все большего числа генов.
Некоторые клетки со временем перестают делиться и погибают, что может быть связано с завершением выполнения определенных функций, как в случае клеток эпидермиса кожи и клеток крови или с повреждением этих клеток факторами окружающей среды, в частности возбудителями болезней. Генетически запрограммированная смерть клетки называется апоптозом, тогда как случайная гибель — некрозом.
Митоз — деление соматических клеток. Фазы митоза
Митоз — способ непрямого деления соматических клеток.
Во время митоза клетка проходит ряд последовательных фаз, в результате которых каждая дочерняя клетка получает такой же набор хромосом, как и в материнской клетке.
Митоз делится на четыре основные фазы: профазу, метафазу, анафазу и телофазу. Профаза — наиболее длительная стадия митоза, в процессе которой происходит конденсация хроматина, в результате чего становятся видны X-образные хромосомы, состоящие из двух хроматид (дочерних хромосом). При этом исчезает ядрышко, центриоли расходятся к полюсам клетки, и начинает формироваться ахроматиновое веретено (веретено деления) из микротрубочек. В конце профазы ядерная оболочка распадается на отдельные пузырьки.
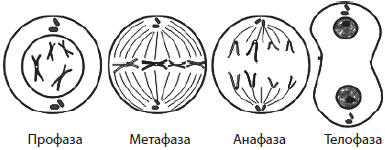
В метафазе хромосомы выстраиваются по экватору клетки своими центромерами, к которым прикрепляются микротрубочки полностью сформированного веретена деления. На этой стадии деления хромосомы наиболее уплотнены и имеют характерную форму, что позволяет изучить кариотип.
В анафазе происходит быстрая репликация ДНК в центромерах, вследствие которой хромосомы расщепляются и хроматиды расходятся к полюсам клетки, растягиваемые микротрубочками. Распределение хроматид должно быть абсолютно равным, поскольку именно этот процесс обеспечивает поддержание постоянства числа хромосом в клетках организма.
На стадии телофазы дочерние хромосомы собираются на полюсах, деспирализуются, вокруг них из пузырьков формируются ядерные оболочки, а во вновь образовавшихся ядрах возникают ядрышки.
После деления ядра происходит деление цитоплазмы — цитокинез, в ходе которого и происходит более или менее равномерное распределение всех органоидов материнской клетки.
Таким образом, в результате митоза из одной материнской клетки образуется две дочерних, каждая из которых является генетической копией материнской (2n2c).
В больных, поврежденных, стареющих клетках и специализированных тканях организма может происходить несколько иной процесс деления — амитоз. Амитозом называют прямое деление эукариотических клеток, при котором не происходит образования генетически равноценных клеток, так как клеточные компоненты распределяются неравномерно. Он встречается у растений в эндосперме, а у животных — в печени, хрящах и роговице глаза.
Мейоз. Фазы мейоза
Мейоз — это способ непрямого деления первичных половых клеток (2n2с), в результате которого образуются гаплоидные клетки (1n1с), чаще всего половые.
В отличие от митоза, мейоз состоит из двух последовательных делений клетки, каждому из которых предшествует интерфаза. Первое деление мейоза (мейоз I) называется редукционным, так как при этом количество хромосом уменьшается вдвое, а второе деление (мейоз II) — эквационным, так как в его процессе количество хромосом сохраняется.
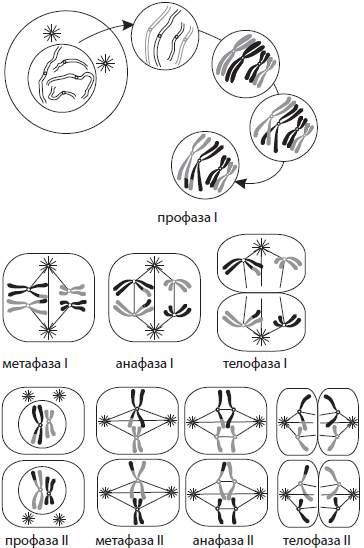
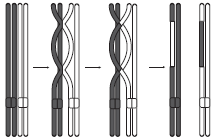
Интерфаза I протекает подобно интерфазе митоза. Мейоз I делится на четыре фазы: профазу I, метафазу I, анафазу I и телофазу I. В профазе I происходят два важнейших процесса — конъюгация и кроссинговер. Конъюгация — это процесс слияния гомологичных (парных) хромосом по всей длине. Образовавшиеся в процессе конъюгации пары хромосом сохраняются до конца метафазы I.
Кроссинговер — взаимный обмен гомологичными участками гомологичных хромосом. В результате кроссинговера хромосомы, полученные организмом от обоих родителей, приобретают новые комбинации генов, что обусловливает появление генетически разнообразного потомства. В конце профазы I, как и в профазе митоза, исчезает ядрышко, центриоли расходятся к полюсам клетки, а ядерная оболочка распадается.
В метафазе I пары хромосом выстраиваются по экватору клетки, к их центромерам прикреп ляются микротрубочки веретена деления.
В анафазе I к полюсам расходятся целые гомологичные хромосомы, состоящие из двух хроматид.
В телофазе I вокруг скоплений хромосом у полюсов клетки образуются ядерные оболочки, формируются ядрышки.
Цитокинез I обеспечивает разделение цитоплазм дочерних клеток.
Образовавшиеся в результате мейоза I дочерние клетки (1n2c) генетически разнородны, поскольку их хромосомы, случайным образом разошедшиеся к полюсам клетки, содержат неодинаковые гены.
Сравнительная характеристика митоза и мейоза
| Признак | Митоз | Мейоз | |
| Какие клетки вступают в деление? | Соматические (2n) | Первичные половые клетки (2n) | |
| Число делений | 1 | 2 | |
| Сколько и каких клеток образуется в процессе деления? | 2 соматические (2n) | 4 половые (n) | |
| Интерфаза | Подготовка клетки к делению, удвоение ДНК | Подготовка клетки к делению, удвоение ДНК | Очень короткая, удвоения ДНК не происходит |
| Фазы | Мейоз I | Мейоз II | |
| Профаза | Конденсация хромосом, исчезновение ядрышка, распад ядерной оболочки | Конденсация хромосом, исчезновение ядрышка, распад ядерной оболочки, могут происходить конъюгация и кроссинговер | Конденсация хромосом, исчезновение ядрышка, распад ядерной оболочки |
| Метафаза | Хромосомы выстраиваются по экватору, формируется веретено деления | По экватору располагаются пары хромосом, формируется веретено деления | Хромосомы выстраиваются по экватору, формируется веретено деления |
| Анафаза | К полюсам расходятся хроматиды | К полюсам расходятся гомологичные хромосомы из двух хроматид | К полюсам расходятся хроматиды |
| Телофаза | Хромосомы деспирализуются, формируются новые ядерные оболочки и ядрышки | Хромосомы деспирализуются, формируются новые ядерные оболочки и ядрышки | Хромосомы деспирализуются, формируются новые ядерные оболочки и ядрышки |
Интерфаза II очень короткая, так как в ней не происходит удвоения ДНК, то есть отсутствует S-период.
Мейоз II также делится на четыре фазы: профазу II, метафазу II, анафазу II и телофазу II. В профазе II протекают те же процессы, что и в профазе I, за исключением конъюгации и кроссинговера.
В метафазе II хромосомы располагаются вдоль экватора клетки.
В анафазе II хромосомы расщепляются в центромерах и к полюсам растягиваются уже хроматиды.
В телофазе II вокруг скоплений дочерних хромосом формируются ядерные оболочки и ядрышки.
После цитокинеза II генетическая формула всех четырех дочерних клеток — 1n1c, однако все они имеют различный набор генов, что является результатом кроссинговера и случайного сочетания хромосом материнского и отцовского организмов в дочерних клетках.
Развитие половых клеток у растений и животных
Гаметогенез (от греч. гамете — жена, гаметес — муж и генезис — происхождение, возникновение) — это процесс образования зрелых половых клеток.
Так как для полового размножения чаще всего необходимы две особи — женская и мужская, продуцирующие различные половые клетки — яйцеклетки и спермии, то и процессы образования этих гамет должны быть различны.
Характер процесса в существенной степени зависит и от того, происходит ли он в растительной или животной клетке, поскольку у растений при образовании гамет происходит только митоз, а у животных — и митоз, и мейоз.
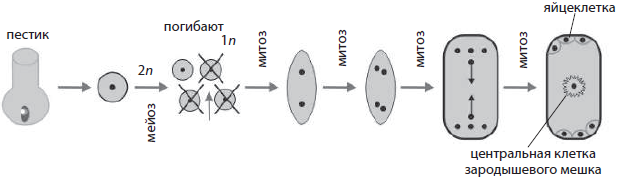
Развитие половых клеток у растений. У покрытосеменных растений образование мужских и женских половых клеток происходит в различных частях цветка — тычинках и пестиках соответственно.
Перед образованием мужских половых клеток — микрогаметогенезом (от греч. микрос — маленький) — происходит микроспорогенез, то есть формирование микроспор в пыльниках тычинок. Этот процесс связан с мейотическим делением материнской клетки, в результате которого возникают четыре гаплоидные микроспоры. Микрогаметогенез сопряжен с митотическим делением микроспоры, дающим мужской гаметофит из двух клеток — крупной вегетативной (сифоногенной) и мелкой генеративной. После деления мужской гаметофит покрывается плотными оболочками и образует пыльцевое зерно. В некоторых случаях еще в процессе созревания пыльцы, а иногда только после переноса на рыльце пестика генеративная клетка делится митотически с образованием двух неподвижных мужских половых клеток — спермиев. Из вегетативной клетки после опыления формируется пыльцевая трубка, по которой спермии проникают в завязь пестика для оплодотворения.
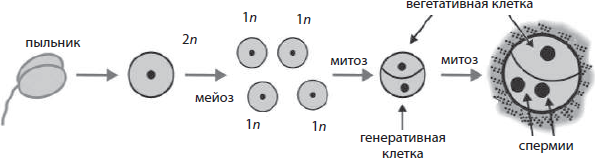
Развитие женских половых клеток у растений называется мегагаметогенезом (от греч. мегас — большой). Он происходит в завязи пестика, чему предшествует мегаспорогенез, в результате которого из материнской клетки мегаспоры, лежащей в нуцеллусе, путем мейотического деления формируются четыре мегаспоры. Одна из мегаспор трижды делится митотически, давая женский гаметофит — зародышевый мешок с восемью ядрами. При последующем обособлении цитоплазм дочерних клеток одна из образовавшихся клеток становится яйцеклеткой, по бокам от которой лежат так называемые синергиды, на противоположном конце зародышевого мешка формируются три антипода, а в центре в результате слияния двух гаплоидных ядер образуется диплоидная центральная клетка.
Развитие половых клеток у животных. У животных различают два процесса образования половых клеток — сперматогенез и овогенез.
Сперматогенез (от греч. сперма, сперматос — семя и генезис — происхождение, возникновение) — это процесс образования зрелых мужских половых клеток — сперматозоидов. У человека он протекает в семенниках, или яичках, и делится на четыре периода: размножение, рост, созревание и формирование.
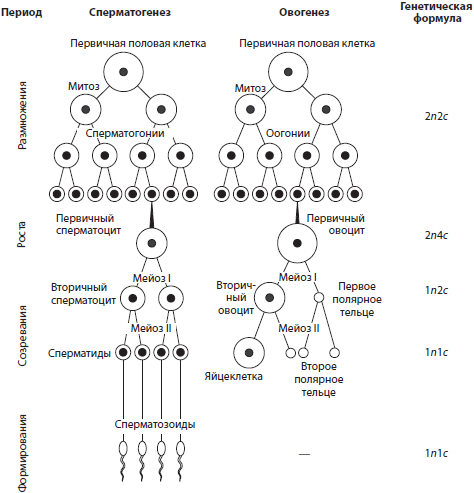
В период размножения первичные половые клетки делятся митотически, вследствие чего образуются диплоидные сперматогонии. В период роста сперматогонии накапливают питательные вещества в цитоплазме, увеличиваются в размерах и превращаются в первичные сперматоциты, или сперматоциты 1-го порядка. Лишь после этого они вступают в мейоз (период созревания), в результате которого образуется сначала два вторичных сперматоцита, или сперматоцита 2-го порядка, а затем — четыре гаплоидных клетки с еще достаточно большим количеством цитоплазмы — сперматиды. В период формирования они утрачивают почти всю цитоплазму и формируют жгутик, превращаясь в сперматозоиды.
Сперматозоиды, или живчики, — очень мелкие подвижные мужские половые клетки, имеющие головку, шейку и хвостик.
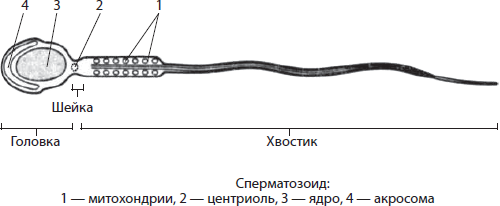
В головке, кроме ядра, находится акросома — видоизмененный комплекс Гольджи, обеспечивающий растворение оболочек яйцеклетки в процессе оплодотворения. В шейке находятся центриоли клеточного центра, а основу хвостика образуют микротрубочки, непосредственно обеспечивающие движение сперматозоида. В нем также расположены митохондрии, обеспечивающие сперматозоид энергией АТФ для движения.
Овогенез (от греч. оон — яйцо и генезис — происхождение, возникновение) — это процесс образования зрелых женских половых клеток — яйцеклеток. У человека он происходит в яичниках и состоит из трех периодов: размножения, роста и созревания. Периоды размножения и роста, аналогичные таковым в сперматогенезе, происходят еще во время внутриутробного развития. При этом из первичных половых клеток в результате митоза образуются диплоидные оогонии, которые превращаются затем в диплоидные первичные ооциты, или ооциты 1-го порядка. Мейоз и последующий цитокинез, протекающие в период созревания, характеризуются неравномерностью деления цитоплазмы материнской клетки, так что в итоге сначала получается один вторичный ооцит, или ооцит 2-го порядка, и первое полярное тельце, а затем из вторичного ооцита — яйцеклетка, сохраняющая весь запас питательных веществ, и второе полярное тельце, тогда как первое полярное тельце делится на два. Полярные тельца забирают избыток генетического материала.
У человека яйцеклетки вырабатываются с промежутком 28–29 суток. Цикл, связанный с созреванием и выходом яйцеклеток, называется менструальным.
Яйцеклетка — крупная женская половая клетка, которая несет не только гаплоидный набор хромосом, но и значительный запас питательных веществ для последующего развития зародыша.
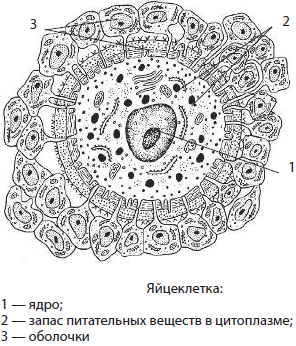
Яйцеклетка у млекопитающих покрыта четырьмя оболочками, снижающими вероятность ее повреждения различными факторами. Диаметр яйцеклетки у человека достигает 150–200 мкм, тогда как у страуса он может составлять несколько сантиметров.
Деление клетки — основа роста, развития и размножения организмов. Роль митоза и мейоза
Если у одноклеточных организмов деление клетки приводит к увеличению количества особей, т. е. размножению, то у многоклеточных этот процесс может иметь различное значение. Так, деление клеток зародыша, начиная с зиготы, является биологической основой взаимосвязанных процессов роста и развития. Подобные же изменения наблюдаются у человека в подростковом возрасте, когда число клеток не только увеличивается, но и происходит качественное изменение организма. В основе размножения многоклеточных организмов также лежит деление клетки, например при бесполом размножении благодаря этому процессу из части организма происходит восстановление целостного, а при половом — в процессе гаметогенеза образуются половые клетки, дающие впоследствии новый организм. Следует отметить, что основные способы деления эукариотической клетки — митоз и мейоз — имеют различное значение в жизненных циклах организмов.
В результате митоза происходит равномерное распределение наследственного материала между дочерними клетками — точными копиями материнской. Без митоза было бы невозможным существование и рост многоклеточных организмов, развивающихся из единственной клетки — зиготы, поскольку все клетки таких организмов должны содержать одинаковую генетическую информацию.
В процессе деления дочерние клетки становятся все более разнообразными по строению и выполняемым функциям, что связано с активацией у них все новых групп генов вследствие межклеточного взаимодействия. Таким образом, митоз необходим для развития организма.
Этот способ деления клеток необходим для процессов бесполого размножения и регенерации (восстановления) поврежденных тканей, а также органов.
Мейоз, в свою очередь, обеспечивает постоянство кариотипа при половом размножении, так как уменьшает вдвое набор хромосом перед половым размножением, который затем восстанавливается в результате оплодотворения. Кроме того, мейоз приводит к появлению новых комбинаций родительских генов благодаря кроссинговеру и случайному сочетанию хромосом в дочерних клетках. Благодаря этому потомство получается генетически разнообразным, что дает материал для естественного отбора и является материальной основой эволюции. Изменение числа, формы и размеров хромосом, с одной стороны, может привести к появлению различных отклонений в развитии организма и даже его гибели, а с другой — может привести к появлению особей, более приспособленных к среде обитания.
Таким образом, клетка является единицей роста, развития и размножения организмов.

Онлайн-школа «Турбо»
- Прямая связь с преподавателем
- Письменные дз с проверкой
- Интересные онлайн-занятия
- Душевное комьюнити