Эволюция: направления, пути, способы эволюции

Вид, его критерии
Основатель современной систематики К. Линней рассматривал вид как группу сходных по морфологическим признакам организмов, которые свободно скрещиваются между собой. По мере развития биологии были получены свидетельства того, что различия между видами намного глубже, и затрагивают химический состав и концентрацию веществ в тканях, направление и скорость химических реакций, характер и интенсивность процессов жизнедеятельности, число и форму хромосом, т. е. вид является наименьшей группой организмов, отражающей их близкое родство. К тому же виды не существуют вечно — они возникают, развиваются, дают начало новым видам и исчезают.
Вид — это совокупность особей, сходных по строению и особенностям процессов жизнедеятельности, имеющих общее происхождение, свободно скрещивающихся между собой в природе и дающих плодовитое потомство.
Все особи одного вида имеют одинаковый кариотип и занимают в природе определенную географическую область — ареал.
Признаки сходства особей одного вида называют критериями вида. Так как ни один из критериев не является абсолютным, для правильного определения вида необходимо использовать совокупность критериев.
Основными критериями вида являются морфологический, физиологический, биохимический, экологический, географический, этологический (поведенческий) и генетический.
- Морфологический — совокупность внешних и внутренних признаков организмов одного вида. Несмотря на то, что у некоторых видов существуют уникальные признаки, с помощью одних только морфологических черт часто очень трудно различить близкородственные виды. Так, в последнее время открыт ряд видов-двойников, обитающих на одной территории, например домовая и курганчиковая мыши, поэтому использовать исключительно морфологический критерий для определения вида недопустимо.
- Физиологический — сходство процессов жизнедеятельности у организмов, в первую очередь, размножения. Он также не является универсальным, поскольку некоторые виды в природе скрещиваются и дают плодовитое потомство.
- Биохимический — сходство химического состава и протекания процессов обмена веществ. Несмотря на то, что данные показатели могут значительно варьировать у разных особей одного вида, в настоящее время им уделяется большое внимание, поскольку особенности строения и состава биополимеров помогают идентифицировать виды даже на молекулярном уровне и устанавливать степень их родства.
- Экологический — различие видов по их принадлежности к определенным экосистемам и экологическим нишам, которые они занимают. Однако многие неродственные виды занимают сходные экологические ниши, поэтому данный критерий может быть использован для выделения вида только в совокупности с другими признаками.
- Географический — существование популяции каждого вида в определенной части биосферы — ареале, который отличается от ареалов всех остальных видов. В связи с тем, что у множества видов границы ареалов совпадают, а также имеется ряд видов-космополитов, ареал которых охватывает огромные пространства, географический критерий также не может служить маркерным «видовым» признаком.
- Генетический — постоянство признаков хромосомного набора — кариотипа — и нуклеотидного состава ДНК у особей одного вида. В связи с тем, что негомологичные хромосомы не могут конъюгировать во время мейоза, потомство от скрещивания особей разных видов с неодинаковым набором хромосом либо не появляется вовсе, либо не плодовито. Это создает репродуктивную изоляцию вида, поддерживает его целостность и обеспечивает реальность существования в природе. Данное правило может нарушаться в случае скрещивания близких по происхождению видов с одинаковым кариотипом или возникновения различных мутаций, однако исключение лишь подтверждает общее правило, и виды следует рассматривать как устойчивые генетические системы. Генетический критерий является основным в системе критериев вида, но также не исчерпывающим.
При всей сложности системы критериев вид нельзя представить как группу абсолютно идентичных по всем параметрам организмов, то есть клонов. Наоборот, для многих видов характерно значительное разнообразие даже внешних признаков, как, например, для одних популяций божьих коровок характерно преобладание в окраске красного цвета, а для других — черного.
Популяция — структурная единица вида и элементарная единица эволюции
Трудно представить, чтобы в реальности особи одного вида были равномерно распределены по земной поверхности в пределах ареала, поскольку, например, лягушка озерная обитает в основном в достаточно редких стоячих пресных водоемах, и вряд ли ее можно встретить на полях и в лесах. Виды в природе чаще всего распадаются на отдельные группы, в зависимости от подходящих по совокупности условий местам обитания — популяции.
Популяция — группа особей одного вида, занимающих часть его ареала, свободно скрещивающихся между собой и относительно обособленных от других совокупностей особей того же вида в течение более или менее длительного времени.
Популяции могут быть разделены не только пространственно, они могут обитать даже на одной территории, но иметь различия в пищевых предпочтениях, сроках размножения и т. д..
Таким образом, вид — это совокупность популяций особей, обладающих рядом общих морфологических, физиологических, биохимических признаков и типов взаимоотношений с окружающей средой, населяющих определенный ареал, а также способных скрещиваться между собой с образованием плодовитого потомства, но почти или совсем не скрещивающихся с другими группами особей того же вида.
Внутри видов с большими ареалами, охватывающими территории с разными условиями жизни, иногда различают и подвиды — большие популяции или группы соседних популяций, имеющих стойкие морфологические отличия от других популяций.
Популяции разбросаны по земной поверхности не случайным образом, они привязаны к конкретным ее участкам. Совокупность всех факторов неживой природы, необходимых для проживания особей данного вида, называется местообитанием. Однако только этих факторов может быть недостаточно для занятия этого участка популяцией, поскольку она должна быть еще вовлечена в тесное взаимодействие с популяциями других видов, то есть занять определенное место в сообществе живых организмов — экологическую нишу. Так, австралийский сумчатый медведь коала при всех прочих равных условиях не может существовать без своего основного источника питания — эвкалипта.
Образующие неразрывное единство в одних и тех же местообитаниях популяции различных видов обыкновенно обеспечивают более или менее замкнутый круговорот веществ и являются элементарными экологическими системами (экосистемами) — биогеоценозами.
При всей своей требовательности к условиям окружающей среды популяции одного вида неоднородны по занимаемой площади, численности, плотности и пространственному размещению особей, часто образующих более мелкие группы (семьи, стаи, стада и др.), полу, возрасту, генофонду и т. д., поэтому различают их размерную, возрастную, половую, пространственную, генетическую, этологическую и другие структуры, а также динамику.
Важными характирестиками популяции являются генофонд — совокупность генов, характерных для особей данной популяции или вида, а также частоты определенных аллелей и генотипов. Разные популяции одного и того же вида изначально имеют неодинаковый генофонд, так как новые территории осваивают особи со случайными, а не специально подобранными генами. Под действием внутренних и внешних факторов генофонд подвергается еще более существенным изменениям: обогащается за счет возникновения мутаций и нового сочетания признаков и обедняется в результате выпадения отдельных аллелей при гибели или миграции некоторого числа особей.
Новые признаки и их сочетания могут быть полезными, нейтральными или вредными, поэтому в популяции выживают и успешно размножаются только приспособленные к данным условиям окружающей среды особи. Однако в двух различных точках земной поверхности условия окружающей среды никогда не бывают полностью идентичными, поэтому и направление изменений даже в двух соседних популяциях может быть совершенно противоположным или они будут протекать с различной скоростью. Результатом изменений генофонда является расхождение популяций по морфологическим, физиологическим, биохимическим и другим признакам. Если популяции при этом еще и изолированы друг от друга, то они могут давать начало новым видам.
Так, возникновение каких-либо препятствий в скрещивании особей различных популяций одного вида, например, вследствие образования горных массивов, изменения русел рек, различий в сроках размножения и т. д., приводит к тому, что популяции постепенно приобретают все больше различий и, в конце концов, становятся различными видами. Некоторое время на границах этих популяций происходит скрещивание особей и возникают гибриды, однако со временем и эти контакты исчезают, т. е. популяции из открытых генетических систем становятся закрытыми.
Несмотря на то, что действию факторов окружающей среды подвергаются в первую очередь отдельные особи, изменение генетического состава у единичного организма является незначительным и проявится в лучшем случае только у его потомков. Подвиды, виды и более крупные таксоны также не подходят на роль элементарных единиц эволюции, поскольку они не отличаются морфологическим, физиологическим, биохимическим, экологическим, географическим и генетическим единством, тогда как популяции как наименьшие структурные единицы вида, накапливающие разнообразие случайных изменений, худшие из которых будут отсеяны, соответствуют этому условию и являются элементарными единицами эволюции.
Микроэволюция
Изменение генетической структуры популяций не всегда приводит к образованию нового вида, а может лишь улучшать приспособление популяции к конкретным условиям среды, однако виды не являются вечными и неизменными — они способны развиваться. Этот процесс необратимого исторического изменения живого называется эволюцией. Первично эволюционные преобразования происходят внутри вида на уровне популяций. В их основе лежат, прежде всего, мутационный процесс и естественный отбор, приводящие к изменению генофонда популяций и вида в целом, или даже к образованию новых видов. Совокупность этих элементарных эволюционных событий называется микроэволюцией.
Популяции характеризуются огромным генетическим разнообразием, которое часто не проявляется фенотипически. Генетическое разнообразие возникает вследствие спонтанного мутагенеза, который происходит непрерывно. Большинство мутаций неблагоприятны для организма и снижают жизнеспособность популяции в целом, но, если они рецессивны, то могут сохраняться в гетерозиготе в течение продолжительного времени. Некоторые мутации, не имеющие приспособительной ценности в данных условиях существования, способны приобрести такую ценность в будущем или при освоении новых экологических ниш, создавая, таким образом, резерв наследственной изменчивости.
Значительное влияние на микроэволюционные процессы оказывают колебания численности особей в популяциях, миграции и катастрофы, а также изоляция популяций и видов.
Новый вид является промежуточным результатом эволюции, но никак не ее итогом, поскольку на этом микроэволюция не прерывается — она продолжается далее. Возникающие новые виды в случае удачного сочетания признаков заселяют новые местообитания, и, в свою очередь, дают начало новым видам. Такие группы близкородственных видов объединяются в роды, семейства и т. д. Эволюционные процессы, происходящие в надвидовых группах, называются уже макроэволюцией. В отличие от макроэволюции, микроэволюция протекает в гораздо более сжатые сроки, тогда как первой требуются десятки и сотни тысяч и миллионов лет, как, например, эволюция человека.
В результате микроэволюции формируется все многообразие видов живых организмов, когдалибо существовавших и ныне живущих на Земле.
Вместе с тем эволюция необратима, и уже исчезнувшие виды никогда не возникают вновь. Появляющиеся виды закрепляют все достигнутое в процессе эволюции, однако это не гарантирует того, что в будущем не появятся новые виды, которые будут иметь более совершенные приспособления к условиям окружающей среды.
Образование новых видов
В широком смысле под образованием новых видов понимается не только отщепление от основного ствола вида нового или распад материнского вида на несколько дочерних, а и общее развитие вида как целостной системы, приводящее к существенным изменениям его морфоструктурной организации. Однако чаще все же видообразование рассматривают как процесс формирования новых видов посредством разветвления «родословного древа» вида.
Принципиальное решение проблемы видообразования было предложено Ч. Дарвином. Согласно его теории расселение особей одного вида приводит к образованию популяций, которые вследствие различий условий окружающей среды вынуждены адаптироваться к ним. Это, в свою очередь, влечет за собой обострение внутривидовой борьбы за существование, направляемой естественным отбором. В настоящее время считается, что борьба за существование вовсе не является обязательным фактором видообразования, наоборот, давление отбора в ряде популяций может снижаться. Различие условий существования способствует возникновению неодинаковых приспособительных изменений в популяциях вида, следствием которого является расхождение признаков и свойств популяций — дивергенция.
Однако накопления различий, даже на генетическом уровне, отнюдь не достаточно для появления нового вида. До тех пор, пока различающиеся по каким-либо признакам популяции не только контактируют, но и способны к скрещиванию с образованием плодовитого потомства, они относятся к одному виду. Лишь невозможность перетока генов из одной группы особей в другую, даже в случае разрушения разделяющих их преград, т. е. скрещивания, означает завершение сложнейшего эволюционного процесса формирования нового вида.
Видообразование является продолжением микроэволюционных процессов. Существует точка зрения, что видообразование не может быть сведено к микроэволюции, оно представляет качественный этап эволюции и осуществляется благодаря другим механизмам.
Способы видообразования
Выделяют два основных способа видообразования: аллопатрическое и симпатрическое.
Аллопатрическое, или географическое видообразование является следствием пространственного разделения популяций физическими преградами (горные хребты, моря и реки) вследствие их возникновения или расселения в новые места обитания (географическая изоляция). Поскольку в данном случае генофонд отделившейся популяции существенно отличается от материнской, да и условия в месте ее обитания не будут совпадать с исходными, со временем это приведет к дивергенции и формированию нового вида. Ярким примером географического видообразования является обнаруженное Ч. Дарвином во время путешествия на корабле «Бигль» разнообразие видов вьюрков на Галапагосских островах вблизи побережья Эквадора. По-видимому, отдельные особи единственного обитающего на южноамериканском континенте вьюрка каким-то образом попали на острова, и, вследствие различия условий (в первую очередь доступности пищи) и географической изоляции, они постепенно эволюционировали, образовав группу родственных видов.
В основе симпатрического, или биологического видообразования лежит какая-либо из форм репродуктивной изоляции, при этом новые виды возникают внутри ареала исходного вида. Обязательным условием симпатрического видообразования является быстрая изоляция образовавшихся форм. Это более быстрый процесс, чем аллопатрическое видообразование, а новые формы похожи на исходные предковые.
Симпатрическое видообразование может быть вызвано быстрыми изменениями хромосомного набора (полиплоидизация) или хромосомными перестройками. Иногда новые виды возникают вследствие гибридизации двух исходных видов, как, например, у домашней сливы, являющейся гибридом терна и алычи. В некоторых случаях симпатрическое видообразование связано с разделением экологических ниш у популяций одного вида в пределах единого ареала или сезонной изоляции — расхождения сроков репродукции у растений (разные виды сосны в Калифорнии пылят в феврале и апреле) и сроков размножения у животных.
Из всего многообразия вновь возникающих видов только немногие, наиболее приспособленные, могут существовать длительное время и дать начало новым видам. Причины гибели большинства видов до сих пор неизвестны, скорее всего это обусловлено резкими изменениями климата, геологическими процессами и вытеснением их более приспособленными организмами. В настоящее время одной из причин гибели значительного числа видов является человек, который истребляет наиболее крупных животных и самые красивые растения, и если в XVII веке этот процесс только начался истреблением последнего тура, то в XXI веке ежечасно исчезает более 10 видов.
Сохранение многообразия видов как основа устойчивости биосферы
Несмотря на то, что на планете, по разным оценкам, обитает 5–10 млн еще не описанных видов организмов, о существовании большинства из них мы никогда не узнаем, поскольку ежечасно с лица Земли исчезает около 50 видов. Исчезновение живых организмов в настоящее время совсем не обязательно связано с их физическим истреблением, чаще это обусловлено разрушением в результате человеческой деятельности их природных мест обитания. Гибель отдельного вида вряд ли способна привести к фатальным последствиям для биосферы, однако уже давно установлено, что вымирание одного вида растений влечет за собой гибель 10–12 видов животных, а это уже представляет угрозу как для существования отдельных биогеоценозов, так и для глобальной экосистемы в целом.
Накопленные за предыдущие десятилетия печальные факты вынудили Международный союз охраны природы и природных ресурсов (МСОП) начать в 1949 году сбор информации о редких и находящихся под угрозой исчезновения видах растений и животных. В 1966 году МСОП издал первую «Красную книгу фактов».
Красная книга — это официальный документ, содержащий регулярно обновляемые данные о состоянии и распространении редких и находящихся под угрозой исчезновения видов растений, животных и грибов.
В этом документе принята пятиступенчатая шкала статуса охраняемого вида, причем к первой ступени охраны относят виды, спасение которых невозможно без осуществления специальных мероприятий, а к пятой — восстановленные виды, состояние которых благодаря принятым мерам не вызывает опасений, но они еще не подлежат промышленному использованию. Разработка такой шкалы позволяет направить первоочередные усилия в области охраны именно на самые редкие виды, такие как амурские тигры.
Помимо международного варианта Красной книги, существуют также национальные и региональные ее варианты. В СССР Красная книга была учреждена в 1974 году, а в Российской Федерации порядок ее ведения регламентируется Федеральными законами «Об охране окружающей среды», «О животном мире» и Постановлением Правительства РФ «О Красной книге Российской Федерации». Сегодня в Красную книгу РФ занесено 610 видов растений, 247 видов животных, 42 вида лишайников и 24 вида грибов. Популяции некоторых из них, в свое время оказавшиеся под угрозой исчезновения (бобр европейский, зубр), уже довольно успешно восстановлены.
Под охрану в России взяты следующие виды животных: русская выхухоль, тарбаган (монгольский сурок), белый медведь, кавказская европейская норка, калан, манул, амурский тигр, леопард, снежный барс, сивуч, морж, тюлени, дельфины, киты, лошадь Пржевальского, кулан, розовый пеликан, фламинго обыкновенный, аист черный, лебедь малый, орел степной, беркут, журавль черный, стерх, дрофа, филин, чайка белая, черепаха средиземноморская, полоз японский, гюрза, жаба камышовая, минога каспийская, все виды осетровых рыб, лосось озерный, жук-олень, шмель необыкновенный, аполлон обыкновенный, рак-богомол, жемчужница обыкновенная и др.
К растениям Красной книги РФ относятся 7 видов подснежников, некоторые виды полыни, женьшень настоящий, 7 видов колокольчиков, дуб зубчатый, пролеска пролесковидная, 11 видов касатиков, рябчик русский, тюльпан Шренка, лотос орехоносный, башмачок венерин настоящий, пион тонколистный, ковыль перистый, первоцвет Юлии, прострел (сон-трава) луговой, красавка белладонна, сосна пицундская, тис ягодный, щитовник китайский, полушник озерный, сфагнум мягкий, филлофора курчавая, хара нитевидная и др.
Редкие грибы представлены трюфелем летним, или русским черным трюфелем, трутовиком лакированным и др.
Охрана редких видов в большинстве случаев связана с запретом их уничтожения, сохранением их самих в искусственно созданной среде обитания (зоопарках), охране их местообитаний и созданием низкотемпературных генетических банков.
Наиболее эффективной мерой охраны редких видов является сохранение их местообитаний, которое достигается организацией сети особо охраняемых заповедных территорий, имеющих, в соответствии с Федеральным законом «Об особо охраняемых природных территориях» (1995), международное, федеральное, региональное или местное значение. К ним относятся государственные природные заповедники, национальные парки, природные парки, государственные природные заказники, памятники природы, дендрологические парки, ботанические сады и др.
Государственный природный заповедник — это полностью изъятый из хозяйственного использования особо охраняемый природный комплекс (земля, водные объекты, недра, растительный и животный мир), имеющий природоохранное, научное, эколого-просветительское значение как образец естественной природной среды, типичные или редкие ландшафты, места сохранения генетического фонда растительного и животного мира.
Заповедники, которые входят в международную систему биосферных резерватов, осуществляющих глобальный экологический мониторинг, имеют статус государственных природных биосферных заповедников. Заповедник является природоохранным, научно-исследовательским и эколого- просветительским учреждением, имеющим целью сохранение и изучение естественного хода природных процессов и явлений, генетического фонда растительного и животного мира, отдельных видов и сообществ растений и животных, типичных и уникальных экологических систем.
В настоящее время в России насчитывается около 100 государственных природных заповедников, 19 из которых носят статус биосферного, в т. ч. Байкальский, Баргузинский, Кавказский, «Кедровая падь», Кроноцкий, Приокско-Террасный и др.
В отличие от заповедников, территории (акватории) национальных парков включают в себя природные комплексы и объекты, имеющие особую экологическую, историческую и эстетическую ценности, и предназначены для использования в природоохранных, просветительских, научных и культурных целях и для регулируемого туризма. Такой статус имеют 39 особо охраняемых природных территорий, в т. ч. Забайкальский и Сочинский национальные парки, а также национальные парки «Куршская коса», «Русский Север», «Шушенский бор» и др.
Природные парки являются природоохранными рекреационными учреждениями, находящимися в ведении субъектов Российской Федерации, территории (акватории) которых включают в себя природные комплексы и объекты, имеющие значительную экологическую и эстетическую ценности, и предназначены для использования в природоохранных, просветительских и рекреационных целях.
Государственными природными заказниками являются территории (акватории), имеющие особое значение для сохранения или восстановления природных комплексов или их компонентов и поддержания экологического баланса.
Развитие эволюционных идей. Значение эволюционной теории Ч. Дарвина. Взаимосвязь движущих сил эволюции. Формы естественного отбора, виды борьбы за существование. Синтетическая теория эволюции. Элементарные факторы эволюции. Исследования С. С. Четверикова. Роль эволюционной теории в формировании современной естественнонаучной картины мира
Развитие эволюционных идей
Все теории происхождения и развития органического мира могут быть сведены к трем основным направлениям: креационизму, трансформизму и эволюционизму. Креационизм — это концепция постоянства видов, рассматривающая многообразие органического мира как результат его творения Богом. Данное направление сформировалось в результате установления господства христианской церкви в Европе, опиравшейся на библейские тексты. Видными представителями креационизма были К. Линней и Ж. Кювье.
«Князь ботаников» К. Линней, открывший и описавший сотни новых видов растений, и создавший первую стройную их систему, тем не менее, доказывал, что общее количество видов организмов неизменно со времени сотворения Земли, то есть они не только не появляются вновь, но и не исчезают. Только к концу жизни он пришел к заключению, что делом рук Божьих являются роды, тогда как виды могут развиваться вследствие приспособления к местным условиям.
Вкладом выдающегося французского зоолога Ж. Кювье (1769–1832) в биологию было опиравшееся на многочисленные данные палеонтологии, сравнительной анатомии и физиологии учение о корреляциях — взаимосвязях частей организма. Благодаря этому стало возможным реконструировать по отдельным частям внешний облик животного. Однако в процессе палеонтологических исследований Ж. Кювье не мог не обратить внимания как на явное изобилие ископаемых форм, так и на резкие изменения групп животных в течение геологической истории. Эти данные послужили отправной точкой для формулировки теории катастроф, согласно которой все или почти все организмы на Земле неоднократно погибали в результате периодических стихийных бедствий, а потом планета вновь заселялась видами, пережившими катастрофу. Последователи Ж. Кювье насчитывали до 27 таких катастроф в истории Земли. Соображения же об эволюции представлялись Ж. Кювье оторванными от реальности.
Противоречия в исходных положениях креационизма, которые становились все более очевидными по мере накопления научных фактов, послужили отправной точкой для формирования другой системы взглядов — трансформизма, признающего реальное существование видов и их историческое развитие. Представители данного течения — Ж. Бюффон, И. Гете, Э. Дарвин и Э. Жоффруа Сент-Илер, будучи не в силах вскрыть истинные причины эволюции, сводили их к приспособлению к условиям окружающей среды и наследованию приобретенных признаков. Корни трансформизма можно найти в работах еще древнегреческих и средневековых философов, которые признавали исторические изменения органического мира. Так, Аристотель высказал идею о единстве природы и постепенности перехода от тел неживой природы к растениям, а от них — к животным — «лестнице природы». Основной причиной изменений живых организмов он считал их внутреннее стремление к совершенству.
Французский естествоиспытатель Ж. Бюффон (1707–1788), главным трудом жизни которого была 36-томная «Естественная история», вопреки представлениям креационистов, раздвинул рамки истории Земли до 80–90 тыс. лет. При этом он констатировал единство растительного и животного мира, а также возможность изменения родственных организмов под воздействием факторов окружающей среды в результате одомашнивания и гибридизации.
Английский врач, философ и поэт Э. Дарвин (1731–1802), дед Ч. Дарвина, полагал, что история органического мира насчитывает миллионы лет, а разнообразие животного мира является результатом смешения нескольких «естественных» групп, влияния внешней среды, упражнения и неупражнения органов, и других факторов.
Единство плана строения групп животных считал одним из главных доказательств развития живого мира Э. Жоффруа Сент-Илер (1772–1844). Однако, в отличие от своих предшественников, он склонялся к тому, что изменение видов обусловлено воздействием факторов среды не на взрослые особи, а на зародыши.
Несмотря на то, что в разгоревшейся в 1831 году дискуссии между Ж. Кювье и Э. Жоффруа Сент-Илером в виде серии докладов в Академии наук, явный перевес остался на стороне первого, именно трансформизм стал предтечей эволюционизма. Эволюционизм (теория эволюции, эволюционное учение) — это система взглядов, признающая развитие природы по определенным законам. Он является теоретической вершиной биологии, которая позволяет объяснять наблюдаемые нами многообразие и сложность живых систем. Однако в связи с тем, что эволюционное учение описывает труднодоступные для наблюдения явления, оно сталкивается со значительными затруднениями. Иногда теорию эволюции называют «дарвинизмом» и отождествляют с учением Ч. Дарвина, что является в корне неверным, поскольку, хотя теория Ч. Дарвина и внесла неоценимый вклад в развитие не только эволюционного учения, но и биологии в целом (а также многих других наук), основы эволюционной теории были заложены другими учеными, она продолжает развиваться и по сей день, а «дарвинизм» во многих аспектах имеет лишь историческое значение.
Создателем первой эволюционной теории — ламаркизма — был французский естествоиспытатель Ж. Б. Ламарк (1744–1829). Движущей силой эволюции он считал внутреннее стремление организмов к совершенству (закон градации), однако приспособление к условиям внешней среды вынуждает их к отклонению от этой магистральной линии. При этом органы, которые интенсивно используются животным в процессе жизнедеятельности, развиваются, а ненужные ему, наоборот, ослабляются и даже могут исчезать (закон упражнения и неупражнения органов). Приобретенные в процессе жизни признаки закрепляются и передаются потомкам. Так, наличие перепонок между пальцами ног у водоплавающих птиц он объяснял попытками их предков передвигаться в водной среде, а длинная шея жирафов, по Ламарку, является следствием того, что их предки старались доставать листья с верхушек деревьев.
Недостатками ламаркизма были теоретичность многих построений, а также допущение вмешательства Творца в эволюцию. В процессе развития биологии выяснилось, что индивидуальные изменения, приобретаемые организмами в процессе жизни, большей частью укладываются в пределы фенотипической изменчивости, а их передача практически невозможна. Например, немецкий зоолог и теоретик эволюционного учения А. Вейсман (1834–1914) на протяжении многих поколений отрезал мышам хвосты и всегда получал в потомстве только хвостатых грызунов. Теория Ж. Б. Ламарка не была принята современниками, но на рубеже веков легла в основу так называемого неоламаркизма.
Значение эволюционной теории Ч. Дарвина
Предпосылками для создания наиболее известной эволюционной теории Ч. Дарвина, или дарвинизма, стали публикация в 1778 году труда английского экономиста Т. Мальтуса «Трактат о народонаселении», работы геолога Ч. Лайеля, формулировка клеточной теории, успехи селекции в Англии и собственные наблюдения Ч. Дарвина (1809–1882), сделанные в годы учебы в Кембридже, в экспедиции в качестве натуралиста на корабле «Бигль» и по ее завершении.
Так, Т. Мальтус утверждал, что население Земли увеличивается в геометрической прогрессии, что существенно превышает возможности планеты по обеспечению его продовольствием и ведет к гибели части потомства. Параллели, проведенные Ч. Дарвиным и его соавтором А. Уоллесом (1823–1913), указывали на то, что и в природе особи размножаются с очень большой скоростью, однако численность популяций остается относительно постоянной. Исследования английского геолога Ч. Лайеля позволили установить, что поверхность Земли далеко не всегда была такой, как в настоящее время, а ее изменения были вызваны воздействием воды, ветра, вулканическими извержениями и деятельностью живых организмов. Самого же Ч. Дарвина еще в студенческие годы поразила крайняя степень изменчивости жуков, а во время путешествия — сходство флоры и фауны континентальной Южной Америки и лежащих вблизи нее Галапагосских островов, и в то же время значительное разнообразие видов, например вьюрков и черепах. Кроме того, в экспедиции он мог наблюдать скелеты гигантских вымерших млекопитающих, сходных с современными броненосцами и ленивцами, что существенно поколебало его веру в сотворение видов.
Основные положения теории эволюции были высказаны Ч. Дарвином в 1859 году на заседании Лондонского Королевского общества, а впоследствии развиты в книгах «Происхождение видов путем естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» (1859), «Изменение домашних животных и культурных растений» (1868), «Происхождение человека и половой отбор» (1871), «Выражение эмоций у человека и животных» (1872) и др.
Сущность разработанной Ч. Дарвиным концепции эволюции можно свести к ряду вытекающих друг из друга положений, имеющих соответствующие доказательства:
Сущность разработанной Ч. Дарвиным концепции эволюции можно свести к ряду вытекающих друг из друга положений, имеющих соответствующие доказательства:
Основные ароморфозы в эволюции растений и животных
Растительные и животные организмы, представляющие различные ветви эволюции органического мира, в процессе исторического развития независимо приобретали определенные черты строения, которые будут охарактеризованы далее.
У растений важнейшими из них являются переход от гаплоидности к диплоидности, независимость от воды в процессе оплодотворения, переход от наружного оплодотворения к внутреннему и возникновение двойного оплодотворения, расчленение тела на органы, развитие проводящей системы, усложнение и усовершенствование тканей, а также специализация опыления с помощью насекомых и распространения семян и плодов.
Переход от гаплоидности к диплоидности сделал растения более устойчивыми к действию факторов окружающей среды вследствие снижения риска проявления рецессивных мутаций. Повидимому, это преобразование коснулось предков сосудистых растений, в число которых не входят моховидные, характеризующиеся преобладанием в жизненном цикле гаметофита.
Главные ароморфозы в эволюции животных связаны с возникновением многоклеточности и все большим расчленением всех систем органов, возникновением прочного скелета, развитием центральной нервной системы, а также общественного поведения в различных группах высокоорганизованных животных, что дало толчок и к прогрессу человека.
Макроэволюция. Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация. Причины биологического прогресса и регресса. Гипотезы возникновения жизни на Земле. Основные ароморфозы в эволюции растений и животных. Усложнение живых организмов в процессе эволюции
Макроэволюция
Образование вида ознаменовывает собой новый виток эволюционного процесса, поскольку особи этого вида, будучи более приспособленными к условиям среды, нежели особи материнского вида, постепенно расселяются на новые территории, и уже в его популяциях играют свою созидательную роль мутагенез, популяционные волны, изоляция и естественный отбор. Со временем эти популяции дают начало новым видам, которые вследствие генетической изоляции имеют гораздо больше признаков сходства между собой, чем с видами того рода, от которого отпочковался видродоначальник, и, таким образом, возникает новый род, затем — новое семейство, отряд (порядок), класс и т. д. Совокупность эволюционных процессов, которые приводят к возникновению надвидовых таксонов (родов, семейств, отрядов, классов и т. п.), называется макроэволюцией. Макроэволюционные процессы как бы обобщают микроэволюционные изменения, происходящие в течение длительного времени, выявляя при этом основные тенденции, направления и закономерности эволюции органического мира, которые не поддаются наблюдению на более низком уровне. До сих пор никаких специфических механизмов макроэволюции не выявлено, поэтому считается, что она осуществляется только посредством микроэволюционных процессов, однако эта позиция постоянно подвергается вполне обоснованной критике.
Возникновение сложной иерархической системы органического мира в значительной степени является результатом неодинаковой скорости эволюционирования различных групп организмов. Так, уже упоминавшийся гинкго двулопастный как бы «законсервировался» на тысячи лет, тогда как достаточно близкие к нему сосны существенно изменились за это время.
Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация
Анализируя историю органического мира, можно заметить, что в определенные промежутки времени господствовали отдельные группы организмов, которые затем клонились к упадку либо исчезали вовсе. Таким образом, можно различить три магистральных направления эволюции: биологический прогресс, биологический регресс и биологическая стабилизация. Значительный вклад в разработку учения о направлениях и путях эволюции внесли русские эволюционисты А. Н. Северцов и И. И. Шмальгаузен.
Биологический прогресс связан с биологическим процветанием группы в целом и характеризует ее эволюционный успех. Он отражает закономерное развитие живой природы от простого к сложному, от более низкой степени организации к более высокой. По А. Н. Северцову, критериями биологического прогресса являются увеличение численности особей данной группы, расширение ее ареала, а также появление и развитие в ее составе групп низшего ранга (преобразование вида в род, рода в семейство и т. д.). В настоящее время биологический прогресс наблюдается у покрытосеменных растений, насекомых, костистых рыб и млекопитающих.
По А. Н. Северцову, биологический прогресс может достигаться вследствие определенных морфофизиологических преобразований организмов, при этом он выделил три основные пути достижения: арогенез, аллогенез и катагенез.
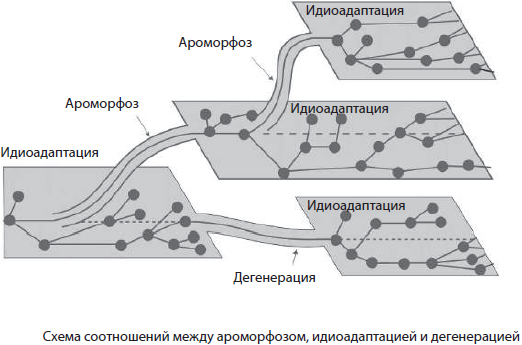
Арогенез, или морфофизиологический прогресс, связан со значительным расширением ареала данной группы организмов вследствие приобретения больших изменений строения — ароморфозов.
Ароморфозом называют эволюционное преобразование строения и функций организма, которое повышает его уровень организации и открывает новые возможности для приспособления к разнообразным условиям существования.
Примерами ароморфозов являются возникновение эукариотической клетки, многоклеточности, появление сердца у рыб и разделение его полной перегородкой у птиц и млекопитающих, формирование цветка у покрытосеменных и т. п.
Аллогенез, в отличие от арогенеза, не сопровождается расширением ареала, однако внутри старого возникает значительное разнообразие форм, имеющих частные приспособления к среде обитания — идиоадаптации.
Идиоадаптация — это мелкое морфофизиологическое приспособление к специальным условиям среды, полезное в борьбе за существование, но не изменяющее уровня организации. Эти изменения иллюстрируют покровительственная окраска у животных, разнообразие ротовых аппаратов у насекомых, колючки растений и др. Не менее удачным примером являются дарвиновы вьюрки, специализирующиеся на различных видах пищи, у которых преобразования вначале затронули клюв, а затем и другие части тела — оперение, хвост и т. п.
Как это ни парадоксально, но и упрощение организации может вести к биологическому прогрессу. Этот путь называется катагенезом.
Дегенерация — это упрощение организмов в процессе эволюции, которое сопровождается потерей определенных функций или органов.
Большей частью она характерна для паразитических организмов и животных, перешедших к прикрепленному способу жизни. Так, растение-паразит повилика европейская лишена хлорофилла, а ее листья редуцированы до чешуек, питается же она за счет поглощения питательных веществ из тела растения-хозяина с помощью специальных присосок. Паразитирующие в тонком кишечнике человека и других животных цепни и лентецы в процессе эволюции принесли свою пищеварительную систему и почти все органы чувств в жертву гипертрофированной половой системе, обеспечивающей необычайную плодовитость (до 11 млрд яиц в течение жизни). Сидячий образ жизни привел у взрослых представителей одной из групп хордовых — асцидий — к исчезновению не только органов чувств и пищеварительной системы, но и самой хорды, обнаруживающейся только у личинок.
Фаза биологического прогресса сменяется фазой биологической стабилизации, сущность которой заключается в сохранении признаков данного вида как наиболее благоприятных в данном микроокружении. По И. И. Шмальгаузену, она вовсе «не означает прекращения эволюции, наоборот, означает максимальную согласованность организма с изменениями среды». В фазе биологической стабилизации находятся «живые ископаемые» латимерия, гингко и др.
Антиподом биологического прогресса является биологический регресс — эволюционный упадок данной группы вследствие невозможности приспособиться к изменениям окружающей среды. Он проявляется в снижении численности популяций, сужении ареалов, уменьшении количества групп низшего ранга в составе высшего таксона. Группе организмов, которая пребывает в состоянии биологического регресса, угрожает вымирание. В истории органического мира можно увидеть много примеров такого явления, и в настоящее время регресс характерен для некоторых папоротников, амфибий и рептилий. С появлением человека биологический регресс зачастую обусловлен его хозяйственной деятельностью.
Направления и пути эволюции органического мира не являются взаимоисключающими, то есть появление ароморфоза не означает, что идиоадаптация или дегенерация произойти уже не может. Напротив, согласно разработанному А. Н. Северцовым и И. И. Шмальгаузеном правилу смены фаз, различные направления эволюционного процесса и пути достижения биологического прогресса закономерно сменяют друг друга. В ходе эволюции эти пути сочетаются: достаточно редкие ароморфозы переводят группу организмов на качественно новый уровень организации, а в дальнейшем историческое развитие идет по пути идиоадаптации либо дегенерации, обеспечивающих приспособление к конкретным условиям среды обитания.
Причины биологического прогресса и регресса
В процессе эволюции планку естественного отбора преодолевают и, соответственно, прогрессируют только те группы организмов, у которых наследственная изменчивость создает достаточное количество комбинаций, которые могут обеспечить выживание группы в целом.
Те же группы, у которых по каким-то причинам не имеется такого резерва, в большинстве случаев обречены на вымирание. Зачастую это связано с низким давлением отбора на предыдущих этапах эволюционного процесса, приведшим к узкой специализации группы или даже дегенеративным явлениям. Следствием этого является невозможность приспособиться к новым условиям среды при ее резких изменениях. Ярким примером тому является внезапная гибель динозавров вследствие падения гигантского небесного тела на Землю 65 млн лет назад, которое повлекло за собой землетрясение, поднятие в воздух миллионов тонн пыли, резкое похолодание, гибель большей части растений и растительноядных животных. В то же время предки современных млекопитающих, не имея узких предпочтений источников питания и будучи теплокровными, смогли пережить эти условия и занять на планете господствующее положение.

Онлайн-школа «Турбо»
- Прямая связь с преподавателем
- Письменные дз с проверкой
- Интересные онлайн-занятия
- Душевное комьюнити