Цитология: деления - 27 линия

Клетка — генетическая единица живого. Хромосомы, их строение (форма и размеры) и функции. Число хромосом и их видовое постоянство. Соматические и половые клетки. Жизненный цикл клетки: интерфаза и митоз. Митоз — деление соматических клеток. Мейоз. Фазы митоза и мейоза. Развитие половых клеток у растений и животных. Деление клетки — основа роста, развития и размножения организмов. Роль мейоза и митоза
Клетка — генетическая единица живого
Несмотря на то, что нуклеиновые кислоты являются носителем генетической информации, реализация этой информации невозможна вне клетки, что легко доказывается на примере вирусов. Данные организмы, содержащие зачастую только ДНК или РНК, не могут самостоятельно воспроизводиться, для этого они должны использовать наследственный аппарат клетки. Даже проникнуть в клетку без помощи самой клетки они не могут, кроме как с использованием механизмов мембранного транспорта или благодаря повреждению клеток. Большинство вирусов нестабильно, они гибнут уже после нескольких часов пребывания на открытом воздухе. Следовательно, клетка является генетической единицей живого, обладающей минимальным набором компонентов для сохранения, изменения и реализации наследственной информации, а также ее передачи потомкам.
Бульшая часть генетической информации эукариотической клетки сосредоточена в ядре. Особенностью ее организации является то, что, в отличие от ДНК прокариотической клетки, молекулы ДНК эукариот не замкнуты и образуют сложные комплексы с белками — хромосомы.
Хромосомы, их строение (форма и размеры) и функции
Хромосома (от греч. хрома — цвет, окраска и сома — тело) — это структура клеточного ядра, которая содержит гены и несет определенную наследственную информацию о признаках и свойствах организма.
Иногда хромосомами называют и кольцевые молекулы ДНК прокариот. Хромосомы способны к самоудвоению, они обладают структурной и функциональной индивидуальностью и сохраняют ее в ряду поколений. Каждая клетка несет всю наследственную информацию организма, но в ней работает только небольшая часть.
Основой хромосомы является двухцепочечная молекула ДНК, упакованная с белками. У эукариот с ДНК взаимодействуют гистоновые и негистоновые белки, тогда как у прокариот гистоновые белки отсутствуют.
Лучше всего хромосомы видны под световым микроскопом в процессе деления клетки, когда они в результате уплотнения приобретают вид палочковидных телец, разделенных первичной перетяжкой — центромерой — на плечи. На хромосоме может быть также и вторичная перетяжка, которая в некоторых случаях отделяет от основной части хромосомы так называемый спутник. Концевые участки хромосом называются теломерами. Теломеры препятствуют слипанию концов хромосом и обеспечивают их прикрепление к оболочке ядра в неделящейся клетке. В начале деления хромосомы удвоены и состоят из двух дочерних хромосом — хроматид, скрепленных в центромере.
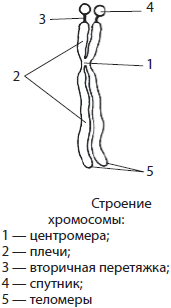
По форме различают равноплечие, неравноплечие и палочковидные хромосомы. Размеры хромосом существенно варьируют, однако средняя хромосома имеет размеры 5 $×$ 1,4 мкм.
В некоторых случаях хромосомы в результате многочисленных удвоений ДНК содержат сотни и тысячи хроматид: такие гигантские хромосомы называются политенными. Они встречаются в слюнных железах личинок дрозофилы, а также в пищеварительных железах аскариды.
Число хромосом и их видовое постоянство. Соматические и половые клетки
Согласно клеточной теории клетка является единицей строения, жизнедеятельности и развития организма. Таким образом, такие важнейшие функции живого, как рост, размножение и развитие организма обеспечиваются на клеточном уровне. Клетки многоклеточных организмов можно разделить на соматические и половые.
Соматические клетки — это все клетки тела, образующиеся в результате митотического деления.
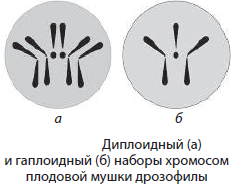
Изучение хромосом позволило установить, что для соматических клеток организма каждого биологического вида характерно постоянное число хромосом. Например, у человека их 46. Набор хромосом соматических клеток называют диплоидным (2n), или двойным.
Половые клетки, или гаметы, — это специализированные клетки, служащие для полового размножения.
В гаметах содержится всегда вдвое меньше хромосом, чем в соматических клетках (у человека — 23), поэтому набор хромосом половых клеток называется гаплоидным (n), или одинарным. Его образование связано с мейотическим делением клетки.
Количество ДНК соматических клеток обозначается как 2c, а половых — 1с. Генетическая формула соматических клеток записывается как 2n2c, а половых — 1n1с.
В ядрах некоторых соматических клеток количество хромосом может отличаться от их количества в соматических клетках. Если это различие больше на один, два, три и т. д. гаплоидных набора, то такие клетки называют полиплоидными (три-, тетра-, пентаплоидными соответственно). В таких клетках процессы метаболизма протекают, как правило, очень интенсивно.
Количество хромосом само по себе не является видоспецифическим признаком, поскольку различные организмы могут иметь равное количество хромосом, а родственные — разное. Например, у малярийного плазмодия и лошадиной аскариды по две хромосомы, а у человека и шимпанзе — 46 и 48 соответственно.
Хромосомы человека делятся на две группы: аутосомы и половые хромосомы (гетерохромосомы). Аутосом в соматических клетках человека насчитывается 22 пары, они одинаковы для мужчин и женщин, а половых хромосом только одна пара, но именно она определяет пол особи. Существует два вида половых хромосом — X и Y. Клетки тела женщины несут по две X-хромосомы, а мужчин — X и Y.
Кариотип — это совокупность признаков хромосомного набора организма (число хромосом, их форма и величина).
Условная запись кариотипа включает общее количество хромосом, половые хромосомы и возможные отклонения в наборе хромосом. Например, кариотип нормального мужчины записывается как 46, XY, а кариотип нормальной женщины — 46, XX.
Жизненный цикл клетки: интерфаза и митоз
Клетки не возникают каждый раз заново, они образуются только в результате деления материнских клеток. После разделения дочерним клеткам требуется некоторое время для формирования органоидов и приобретения соответствующей структуры, которая обеспечила бы выполнение определенной функции. Этот отрезок времени называется созреванием.
Промежуток времени от появления клетки в результате деления до ее разделения или гибели называется жизненным циклом клетки.
У эукариотических клеток жизненный цикл делится на две основные стадии: интерфазу и митоз.
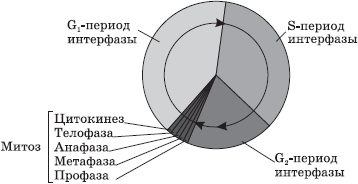
Интерфаза — это промежуток времени в жизненном цикле, в который клетка не делится и нормально функционирует. Интерфаза делится на три периода: G1-, S- и G2-периоды.
G1-период (пресинтетический, постмитотический) — это период роста и развития клетки, в который происходит активный синтез РНК, белков и других веществ, необходимых для полного жизнеобеспечения вновь образовавшейся клетки. К концу этого периода клетка может начать готовиться к удвоению ДНК.
В S-периоде (синтетическом) происходит сам процесс репликации ДНК. Единственным участком хромосомы, который не подвергается репликации, является центромера, поэтому образовавшиеся молекулы ДНК не расходятся полностью, а остаются скрепленными в ней, и в начале деления хромосома имеет X-образный вид. Генетическая формула клетки после удвоения ДНК — 2n4c. Также в S-периоде происходит удвоение центриолей клеточного центра.
G2-период (постсинтетический, премитотический) характеризуется интенсивным синтезом РНК, белков и АТФ, необходимых для процесса деления клетки, а также разделением центриолей, митохондрий и пластид. До конца интерфазы хроматин и ядрышко остаются хорошо различимыми, целостность ядерной оболочки не нарушается, а органоиды не изменяются.
Часть клеток организма способна выполнять свои функции в течение всей жизни организма (нейроны нашего головного мозга, мышечные клетки сердца), а другие существуют непродолжительное время, после чего погибают (клетки кишечного эпителия, клетки эпидермиса кожи). Следовательно, в организме должны постоянно происходить процессы деления клеток и образования новых, которые замещали бы отмершие. Клетки, способные к делению, называют стволовыми. В организме человека они находятся в красном костном мозге, в глубоких слоях эпидермиса кожи и других местах. Используя эти клетки, можно вырастить новый орган, добиться омоложения, а также клонировать организм. Перспективы использования стволовых клеток совершенно ясны, однако морально-этические аспекты этой проблемы все еще обсуждаются, поскольку в большинстве случаев используются эмбриональные стволовые клетки, полученные из убитых при аборте зародышей человека.
Продолжительность интерфазы в клетках растений и животных составляет в среднем 10– 20 часов, тогда как митоз занимает около 1–2 часов.
В ходе последовательных делений в многоклеточных организмах дочерние клетки становятся все более разнообразными, поскольку в них происходит считывание информации со все большего числа генов.
Некоторые клетки со временем перестают делиться и погибают, что может быть связано с завершением выполнения определенных функций, как в случае клеток эпидермиса кожи и клеток крови или с повреждением этих клеток факторами окружающей среды, в частности возбудителями болезней. Генетически запрограммированная смерть клетки называется апоптозом, тогда как случайная гибель — некрозом.
Митоз — деление соматических клеток. Фазы митоза
Митоз — способ непрямого деления соматических клеток.
Во время митоза клетка проходит ряд последовательных фаз, в результате которых каждая дочерняя клетка получает такой же набор хромосом, как и в материнской клетке.
Митоз делится на четыре основные фазы: профазу, метафазу, анафазу и телофазу. Профаза — наиболее длительная стадия митоза, в процессе которой происходит конденсация хроматина, в результате чего становятся видны X-образные хромосомы, состоящие из двух хроматид (дочерних хромосом). При этом исчезает ядрышко, центриоли расходятся к полюсам клетки, и начинает формироваться ахроматиновое веретено (веретено деления) из микротрубочек. В конце профазы ядерная оболочка распадается на отдельные пузырьки.
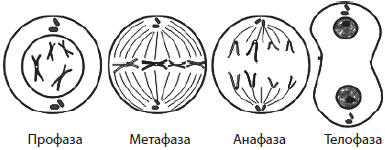
В метафазе хромосомы выстраиваются по экватору клетки своими центромерами, к которым прикрепляются микротрубочки полностью сформированного веретена деления. На этой стадии деления хромосомы наиболее уплотнены и имеют характерную форму, что позволяет изучить кариотип.
В анафазе происходит быстрая репликация ДНК в центромерах, вследствие которой хромосомы расщепляются и хроматиды расходятся к полюсам клетки, растягиваемые микротрубочками. Распределение хроматид должно быть абсолютно равным, поскольку именно этот процесс обеспечивает поддержание постоянства числа хромосом в клетках организма.
На стадии телофазы дочерние хромосомы собираются на полюсах, деспирализуются, вокруг них из пузырьков формируются ядерные оболочки, а во вновь образовавшихся ядрах возникают ядрышки.
После деления ядра происходит деление цитоплазмы — цитокинез, в ходе которого и происходит более или менее равномерное распределение всех органоидов материнской клетки.
Таким образом, в результате митоза из одной материнской клетки образуется две дочерних, каждая из которых является генетической копией материнской (2n2c).
В больных, поврежденных, стареющих клетках и специализированных тканях организма может происходить несколько иной процесс деления — амитоз. Амитозом называют прямое деление эукариотических клеток, при котором не происходит образования генетически равноценных клеток, так как клеточные компоненты распределяются неравномерно. Он встречается у растений в эндосперме, а у животных — в печени, хрящах и роговице глаза.
Мейоз. Фазы мейоза
Мейоз — это способ непрямого деления первичных половых клеток (2n2с), в результате которого образуются гаплоидные клетки (1n1с), чаще всего половые.
В отличие от митоза, мейоз состоит из двух последовательных делений клетки, каждому из которых предшествует интерфаза. Первое деление мейоза (мейоз I) называется редукционным, так как при этом количество хромосом уменьшается вдвое, а второе деление (мейоз II) — эквационным, так как в его процессе количество хромосом сохраняется.
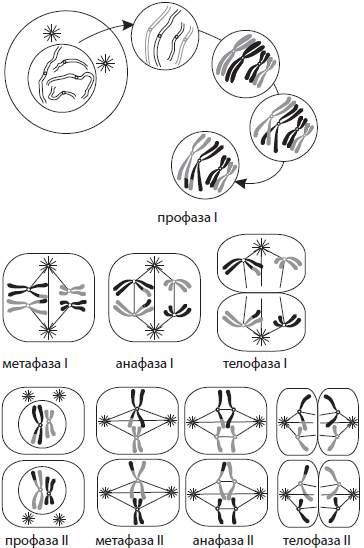
Интерфаза I протекает подобно интерфазе митоза. Мейоз I делится на четыре фазы: профазу I, метафазу I, анафазу I и телофазу I. В профазе I происходят два важнейших процесса — конъюгация и кроссинговер. Конъюгация — это процесс слияния гомологичных (парных) хромосом по всей длине. Образовавшиеся в процессе конъюгации пары хромосом сохраняются до конца метафазы I.
Кроссинговер — взаимный обмен гомологичными участками гомологичных хромосом. В результате кроссинговера хромосомы, полученные организмом от обоих родителей, приобретают новые комбинации генов, что обусловливает появление генетически разнообразного потомства. В конце профазы I, как и в профазе митоза, исчезает ядрышко, центриоли расходятся к полюсам клетки, а ядерная оболочка распадается.
В метафазе I пары хромосом выстраиваются по экватору клетки, к их центромерам прикреп ляются микротрубочки веретена деления.
В анафазе I к полюсам расходятся целые гомологичные хромосомы, состоящие из двух хроматид.
В телофазе I вокруг скоплений хромосом у полюсов клетки образуются ядерные оболочки, формируются ядрышки.
Цитокинез I обеспечивает разделение цитоплазм дочерних клеток.
Образовавшиеся в результате мейоза I дочерние клетки (1n2c) генетически разнородны, поскольку их хромосомы, случайным образом разошедшиеся к полюсам клетки, содержат неодинаковые гены.
Сравнительная характеристика митоза и мейоза
| Признак | Митоз | Мейоз | |
| Какие клетки вступают в деление? | Соматические (2n) | Первичные половые клетки (2n) | |
| Число делений | 1 | 2 | |
| Сколько и каких клеток образуется в процессе деления? | 2 соматические (2n) | 4 половые (n) | |
| Интерфаза | Подготовка клетки к делению, удвоение ДНК | Подготовка клетки к делению, удвоение ДНК | Очень короткая, удвоения ДНК не происходит |
| Фазы | Мейоз I | Мейоз II | |
| Профаза | Конденсация хромосом, исчезновение ядрышка, распад ядерной оболочки | Конденсация хромосом, исчезновение ядрышка, распад ядерной оболочки, могут происходить конъюгация и кроссинговер | Конденсация хромосом, исчезновение ядрышка, распад ядерной оболочки |
| Метафаза | Хромосомы выстраиваются по экватору, формируется веретено деления | По экватору располагаются пары хромосом, формируется веретено деления | Хромосомы выстраиваются по экватору, формируется веретено деления |
| Анафаза | К полюсам расходятся хроматиды | К полюсам расходятся гомологичные хромосомы из двух хроматид | К полюсам расходятся хроматиды |
| Телофаза | Хромосомы деспирализуются, формируются новые ядерные оболочки и ядрышки | Хромосомы деспирализуются, формируются новые ядерные оболочки и ядрышки | Хромосомы деспирализуются, формируются новые ядерные оболочки и ядрышки |
Интерфаза II очень короткая, так как в ней не происходит удвоения ДНК, то есть отсутствует S-период.
Мейоз II также делится на четыре фазы: профазу II, метафазу II, анафазу II и телофазу II. В профазе II протекают те же процессы, что и в профазе I, за исключением конъюгации и кроссинговера.
В метафазе II хромосомы располагаются вдоль экватора клетки.
В анафазе II хромосомы расщепляются в центромерах и к полюсам растягиваются уже хроматиды.
В телофазе II вокруг скоплений дочерних хромосом формируются ядерные оболочки и ядрышки.
После цитокинеза II генетическая формула всех четырех дочерних клеток — 1n1c, однако все они имеют различный набор генов, что является результатом кроссинговера и случайного сочетания хромосом материнского и отцовского организмов в дочерних клетках.
Развитие половых клеток у растений и животных
Гаметогенез (от греч. гамете — жена, гаметес — муж и генезис — происхождение, возникновение) — это процесс образования зрелых половых клеток.
Так как для полового размножения чаще всего необходимы две особи — женская и мужская, продуцирующие различные половые клетки — яйцеклетки и спермии, то и процессы образования этих гамет должны быть различны.
Характер процесса в существенной степени зависит и от того, происходит ли он в растительной или животной клетке, поскольку у растений при образовании гамет происходит только митоз, а у животных — и митоз, и мейоз.
Развитие половых клеток у растений. У покрытосеменных растений образование мужских и женских половых клеток происходит в различных частях цветка — тычинках и пестиках соответственно.
Перед образованием мужских половых клеток — микрогаметогенезом (от греч. микрос — маленький) — происходит микроспорогенез, то есть формирование микроспор в пыльниках тычинок. Этот процесс связан с мейотическим делением материнской клетки, в результате которого возникают четыре гаплоидные микроспоры. Микрогаметогенез сопряжен с митотическим делением микроспоры, дающим мужской гаметофит из двух клеток — крупной вегетативной (сифоногенной) и мелкой генеративной. После деления мужской гаметофит покрывается плотными оболочками и образует пыльцевое зерно. В некоторых случаях еще в процессе созревания пыльцы, а иногда только после переноса на рыльце пестика генеративная клетка делится митотически с образованием двух неподвижных мужских половых клеток — спермиев. Из вегетативной клетки после опыления формируется пыльцевая трубка, по которой спермии проникают в завязь пестика для оплодотворения.
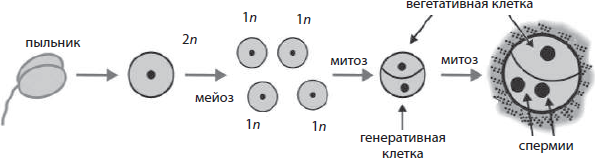
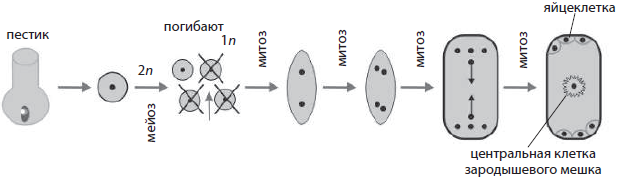
Развитие женских половых клеток у растений называется мегагаметогенезом (от греч. мегас — большой). Он происходит в завязи пестика, чему предшествует мегаспорогенез, в результате которого из материнской клетки мегаспоры, лежащей в нуцеллусе, путем мейотического деления формируются четыре мегаспоры. Одна из мегаспор трижды делится митотически, давая женский гаметофит — зародышевый мешок с восемью ядрами. При последующем обособлении цитоплазм дочерних клеток одна из образовавшихся клеток становится яйцеклеткой, по бокам от которой лежат так называемые синергиды, на противоположном конце зародышевого мешка формируются три антипода, а в центре в результате слияния двух гаплоидных ядер образуется диплоидная центральная клетка.
Развитие половых клеток у животных. У животных различают два процесса образования половых клеток — сперматогенез и овогенез.
Сперматогенез (от греч. сперма, сперматос — семя и генезис — происхождение, возникновение) — это процесс образования зрелых мужских половых клеток — сперматозоидов. У человека он протекает в семенниках, или яичках, и делится на четыре периода: размножение, рост, созревание и формирование.
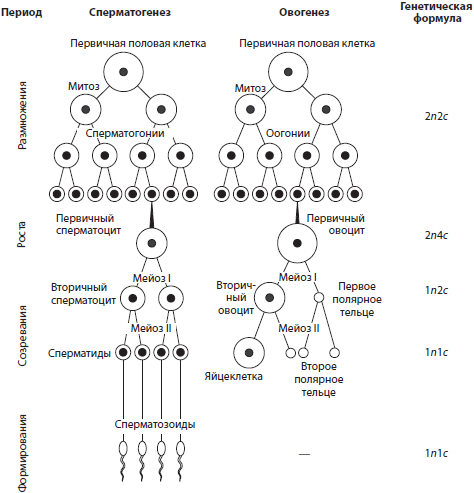
В период размножения первичные половые клетки делятся митотически, вследствие чего образуются диплоидные сперматогонии. В период роста сперматогонии накапливают питательные вещества в цитоплазме, увеличиваются в размерах и превращаются в первичные сперматоциты, или сперматоциты 1-го порядка. Лишь после этого они вступают в мейоз (период созревания), в результате которого образуется сначала два вторичных сперматоцита, или сперматоцита 2-го порядка, а затем — четыре гаплоидных клетки с еще достаточно большим количеством цитоплазмы — сперматиды. В период формирования они утрачивают почти всю цитоплазму и формируют жгутик, превращаясь в сперматозоиды.
Сперматозоиды, или живчики, — очень мелкие подвижные мужские половые клетки, имеющие головку, шейку и хвостик.
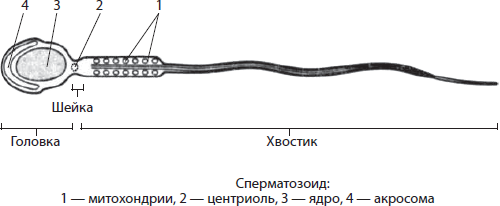
В головке, кроме ядра, находится акросома — видоизмененный комплекс Гольджи, обеспечивающий растворение оболочек яйцеклетки в процессе оплодотворения. В шейке находятся центриоли клеточного центра, а основу хвостика образуют микротрубочки, непосредственно обеспечивающие движение сперматозоида. В нем также расположены митохондрии, обеспечивающие сперматозоид энергией АТФ для движения.
Овогенез (от греч. оон — яйцо и генезис — происхождение, возникновение) — это процесс образования зрелых женских половых клеток — яйцеклеток. У человека он происходит в яичниках и состоит из трех периодов: размножения, роста и созревания. Периоды размножения и роста, аналогичные таковым в сперматогенезе, происходят еще во время внутриутробного развития. При этом из первичных половых клеток в результате митоза образуются диплоидные оогонии, которые превращаются затем в диплоидные первичные ооциты, или ооциты 1-го порядка. Мейоз и последующий цитокинез, протекающие в период созревания, характеризуются неравномерностью деления цитоплазмы материнской клетки, так что в итоге сначала получается один вторичный ооцит, или ооцит 2-го порядка, и первое полярное тельце, а затем из вторичного ооцита — яйцеклетка, сохраняющая весь запас питательных веществ, и второе полярное тельце, тогда как первое полярное тельце делится на два. Полярные тельца забирают избыток генетического материала.
У человека яйцеклетки вырабатываются с промежутком 28–29 суток. Цикл, связанный с созреванием и выходом яйцеклеток, называется менструальным.
Яйцеклетка — крупная женская половая клетка, которая несет не только гаплоидный набор хромосом, но и значительный запас питательных веществ для последующего развития зародыша.
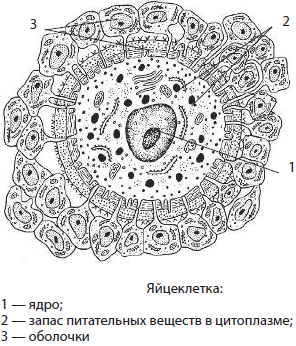
Яйцеклетка у млекопитающих покрыта четырьмя оболочками, снижающими вероятность ее повреждения различными факторами. Диаметр яйцеклетки у человека достигает 150–200 мкм, тогда как у страуса он может составлять несколько сантиметров.
Деление клетки — основа роста, развития и размножения организмов. Роль митоза и мейоза
Если у одноклеточных организмов деление клетки приводит к увеличению количества особей, т. е. размножению, то у многоклеточных этот процесс может иметь различное значение. Так, деление клеток зародыша, начиная с зиготы, является биологической основой взаимосвязанных процессов роста и развития. Подобные же изменения наблюдаются у человека в подростковом возрасте, когда число клеток не только увеличивается, но и происходит качественное изменение организма. В основе размножения многоклеточных организмов также лежит деление клетки, например при бесполом размножении благодаря этому процессу из части организма происходит восстановление целостного, а при половом — в процессе гаметогенеза образуются половые клетки, дающие впоследствии новый организм. Следует отметить, что основные способы деления эукариотической клетки — митоз и мейоз — имеют различное значение в жизненных циклах организмов.
В результате митоза происходит равномерное распределение наследственного материала между дочерними клетками — точными копиями материнской. Без митоза было бы невозможным существование и рост многоклеточных организмов, развивающихся из единственной клетки — зиготы, поскольку все клетки таких организмов должны содержать одинаковую генетическую информацию.
В процессе деления дочерние клетки становятся все более разнообразными по строению и выполняемым функциям, что связано с активацией у них все новых групп генов вследствие межклеточного взаимодействия. Таким образом, митоз необходим для развития организма.
Этот способ деления клеток необходим для процессов бесполого размножения и регенерации (восстановления) поврежденных тканей, а также органов.
Мейоз, в свою очередь, обеспечивает постоянство кариотипа при половом размножении, так как уменьшает вдвое набор хромосом перед половым размножением, который затем восстанавливается в результате оплодотворения. Кроме того, мейоз приводит к появлению новых комбинаций родительских генов благодаря кроссинговеру и случайному сочетанию хромосом в дочерних клетках. Благодаря этому потомство получается генетически разнообразным, что дает материал для естественного отбора и является материальной основой эволюции. Изменение числа, формы и размеров хромосом, с одной стороны, может привести к появлению различных отклонений в развитии организма и даже его гибели, а с другой — может привести к появлению особей, более приспособленных к среде обитания.
Таким образом, клетка является единицей роста, развития и размножения организмов.
Обмен веществ и превращения энергии — свойства живых организмов. Энергетический и пластический обмен, их взаимосвязь. Стадии энергетического обмена. Брожение и дыхание. Фотосинтез, его значение, космическая роль. Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь. Хемосинтез. Роль хемосинтезирующих бактерий на Земле
Обмен веществ и превращения энергии — свойства живых организмов
Клетку можно уподобить миниатюрной химической фабрике, на которой происходят сотни и тысячи химических реакций.
Обмен веществ — совокупность химических превращений, направленных на сохранение и самовоспроизведение биологических систем.
Он включает в себя поступление веществ в организм в процессе питания и дыхания, внутриклеточный обмен веществ, или метаболизм, а также выделение конечных продуктов обмена.
Обмен веществ неразрывно связан с процессами превращения одних видов энергии в другие. Например, в процессе фотосинтеза световая энергия запасается в виде энергии химических связей сложных органических молекул, а в процессе дыхания она высвобождается и расходуется на синтез новых молекул, механическую и осмотическую работу, рассеивается в виде тепла и т. д.
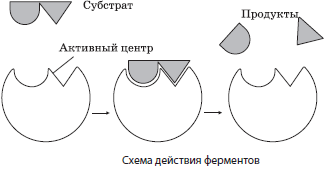
Протекание химических реакций в живых организмах обеспечивается благодаря биологическим катализаторам белковой природы — ферментам, или энзимам. Как и другие катализаторы, ферменты ускоряют протекание химических реакций в клетке в десятки и сотни тысяч раз, а иногда и вообще делают их возможными, но не изменяют при этом ни природы, ни свойств конечного продукта (продуктов) реакции и не изменяются сами. Ферменты могут быть как простыми, так и сложными белками, в состав которых, кроме белковой части, входит и небелковая — кофактор (кофермент). Примерами ферментов являются амилаза слюны, расщепляющая полисахариды при длительном пережевывании, и пепсин, обеспечивающий переваривание белков в желудке.
Ферменты отличаются от катализаторов небелковой природы высокой специфичностью действия, значительным увеличением с их помощью скорости реакции, а также возможностью регуляции действия за счет изменения условий протекания реакции либо взаимодействия с ними различных веществ. К тому же и условия, в которых протекает ферментный катализ, существенно отличаются от тех, при которых идет неферментный: оптимальной для функционирования ферментов в организме человека является температура $37°С$, давление должно быть близким к атмосферному, а $рН$ среды может существенно колебаться. Так, для амилазы необходима щелочная среда, а для пепсина — кислая.
Механизм действия ферментов заключается в снижении энергии активации веществ (субстратов), вступающих в реакцию, за счет образования промежуточных фермент-субстратных комплексов.
Энергетический и пластический обмен, их взаимосвязь
Метаболизм складывается из двух одновременно протекающих в клетке процессов: пластического и энергетического обменов.
Пластический обмен (анаболизм, ассимиляция) представляет собой совокупность реакций синтеза, которые идут с затратой энергии АТФ. В процессе пластического обмена синтезируются органические вещества, необходимые клетке. Примером реакций пластического обмена являются фотосинтез, биосинтез белка и репликация (самоудвоение) ДНК.
Энергетический обмен (катаболизм, диссимиляция) — это совокупность реакций расщепления сложных веществ до более простых. В результате энергетического обмена выделяется энергия, запасаемая в виде АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.
Пластический и энергетический обмены неразрывно связаны, поскольку в процессе пластического обмена синтезируются органические вещества и для этого необходима энергия АТФ, а в процессе энергетического обмена органические вещества расщепляются и высвобождается энергия, которая затем будет израсходована на процессы синтеза.
Энергию организмы получают в процессе питания, а высвобождают ее и переводят в доступную форму в основном в процессе дыхания. По способу питания все организмы делятся на автотрофов и гетеротрофов. Автотрофы способны самостоятельно синтезировать органические вещества из неорганических, а гетеротрофы используют исключительно готовые органические вещества.
Стадии энергетического обмена
Несмотря на всю сложность реакций энергетического обмена, его условно подразделяют на три этапа: подготовительный, анаэробный (бескислородный) и аэробный (кислородный).
На подготовительном этапе молекулы полисахаридов, липидов, белков, нуклеиновых кислот распадаются на более простые, например, глюкозу, глицерин и жирные кислоты, аминокислоты, нуклеотиды и др. Этот этап может протекать непосредственно в клетках либо в кишечнике, откуда расщепленные вещества доставляются с током крови.
Анаэробный этап энергетического обмена сопровождается дальнейшим расщеплением мономеров органических соединений до еще более простых промежуточных продуктов, например, пировиноградной кислоты, или пирувата. Он не требует присутствия кислорода, и для многих организмов, обитающих в иле болот или в кишечнике человека, является единственным способом получения энергии. Анаэробный этап энергетического обмена протекает в цитоплазме.
Бескислородному расщеплению могут подвергаться различные вещества, однако довольно часто субстратом реакций оказывается глюкоза. Процесс ее бескислородного расщепления называется гликолизом. При гликолизе молекула глюкозы теряет четыре атома водорода, т. е. окисляется, при этом образуются две молекулы пировиноградной кислоты, две молекулы АТФ и две молекулы восстановленного переносчика водорода $НАДН + Н^{+}$:
$С_6Н_{12}О_6 + 2Н_3РО_4 + 2АДФ + 2НАД → 2С_3Н_4О_3 + 2АТФ + 2НАДН + Н^{+} + 2Н_2О$.
Образование АТФ из АДФ происходит вследствие прямого переноса фосфат-аниона с предварительно фосфорилированного сахара и называется субстратным фосфорилированием.
Аэробный этап энергетического обмена может происходить только в присутствии кислорода, при этом промежуточные соединения, образовавшиеся в процессе бескислородного расщепления, окисляются до конечных продуктов (углекислого газа и воды) и выделяется большая часть энергии, запасенной в химических связях органических соединений. Она переходит в энергию макроэргических связей 36 молекул АТФ. Этот этап также называется тканевым дыханием. В случае отсутствия кислорода промежуточные соединения превращаются в другие органические вещества, и этот процесс называется брожением.
Дыхание
Механизм клеточного дыхания схематически изображен на рис.
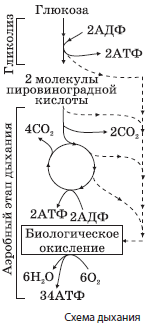
Аэробное дыхание происходит в митохондриях, при этом пировиноградная кислота сначала утрачивает один атом углерода, что сопровождается синтезом одного восстановительного эквивалента $НАДН + Н^{+}$ и молекулы ацетилкофермента А (ацетил-КоА):
$С_3Н_4О_3 + НАД + Н~КоА → СН_3СО~КоА + НАДН + Н^{+} + СО_2↑$.
Ацетил-КоА в матриксе митохондрий вовлекается в цепь химических реакций, совокупность которых называется циклом Кребса (циклом трикарбоновых кислот, циклом лимонной кислоты). В ходе этих превращений образуется две молекулы АТФ, ацетил-КоА полностью окисляется до углекислого газа, а его ионы водорода и электроны присоединяются к переносчикам водорода $НАДН + Н^{+}$ и $ФАДН_2$. Переносчики транспортируют протоны водорода и электроны к внутренним мембранам митохондрий, образующим кристы. При помощи белков-переносчиков протоны водорода нагнетаются в межмембранное пространство, а электроны передаются по так называемой дыхательной цепи ферментов, расположенной на внутренней мембране митохондрий, и сбрасываются на атомы кислорода:
$O_2+2e^{-}→O_2^-$.
Следует отметить, что некоторые белки дыхательной цепи содержат железо и серу.
Из межмембранного пространства протоны водорода транспортируются обратно в матрикс митохондрий с помощью специальных ферментов — АТФ-синтаз, а выделяющаяся при этом энергия расходуется на синтез 34 молекул АТФ из каждой молекулы глюкозы. Этот процесс называется окислительным фосфорилированием. В матриксе митохондрий протоны водорода реагируют с радикалами кислорода с образованием воды:
$4H^{+}+O_2^-→2H_2O$.
Совокупность реакций кислородного дыхания может быть выражена следующим образом:
$2С_3Н_4О_3 + 6О_2 + 36Н_3РО_4 + 36АДФ → 6СО_2↑ + 38Н_2О + 36АТФ.$
Суммарное уравнение дыхания выглядит таким образом:
$С_6Н_{12}О_6 + 6О_2 + 38Н_3РО_4 + 38АДФ → 6СО_2↑ + 40Н_2О + 38АТФ.$
Брожение
В отсутствие кислорода или при его недостатке происходит брожение. Брожение является эволюционно более ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку в результате брожения образуются органические вещества, все еще богатые энергией. Различают несколько основных видов брожения: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода в ходе брожения пировиноградная кислота восстанавливается до молочной кислоты, при этом образовавшиеся ранее восстановительные эквиваленты расходуются, и остаются всего две молекулы АТФ:
$2С_3Н_4О_3 + 2НАДН + Н^{+} → 2С_3Н_6О_3 + 2НАД$.
При брожении с помощью дрожжевых грибов пировиноградная кислота в присутствии кислорода превращается в этиловый спирт и оксид углерода (IV):
$С_3Н_4О_3 + НАДН + Н^{+} → С_2Н_5ОН + СО_2↑ + НАД^{+}$.
При брожении с помощью микроорганизмов из пировиноградной кислоты могут образоваться также уксусная, масляная, муравьиная кислоты и др.
АТФ, полученная в результате энергетического обмена, расходуется в клетке на различные виды работы: химическую, осмотическую, электрическую, механическую и регуляторную. Химическая работа заключается в биосинтезе белков, липидов, углеводов, нуклеиновых кислот и других жизненно важных соединений. К осмотической работе относят процессы поглощения клеткой и выведения из нее веществ, которые во внеклеточном пространстве находятся в концентрациях, больших, чем в самой клетке. Электрическая работа тесно взаимосвязана с осмотической, поскольку именно в результате перемещения заряженных частиц через мембраны формируется заряд мембраны и приобретаются свойства возбудимости и проводимости. Механическая работа сопряжена с движением веществ и структур внутри клетки, а также клетки в целом. К регуляторной работе относят все процессы, направленные на координацию процессов в клетке.
Половое размножение происходит с образованием половых клеток — гамет — и последующим оплодотворением. Пыльцевое зерно у цветковых растений представляет собой сильно редуцированный мужской гаметофит, содержащий две клетки: вегетативную и генеративную. Генеративная клетка после созревания пыльцевого зерна делится на две мужские половые клетки — спермия. Особенностью спермиев цветковых и голосеменных растений является их неспособность к активному передвижению.
Женские половые клетки растений — яйцеклетки — формируются внутри семязачатков. Семязачаток — это многоклеточное образование семенных растений, из которого затем сформируется семя. Он покрыт плотными покровами — интегументом — со специальным отверстием — пыльцевходом, или микропиле. В семязачатке формируется редуцированный женский гаметофит — зародышевый мешок, образованный шестью гаплоидными (яйцеклеткой, синергидами и антиподами) и одной диплоидной (центральной) клетками. Прямо у пыльцевхода обычно лежат яйцеклетка и сопровождающие ее две синергиды. Функция синергид состоит в том, чтобы выделять химические вещества, на которые ориентируется прорастающая пыльцевая трубка. На противоположном полюсе зародышевого мешка располагаются три антипода, тогда как центральная клетка находится посередине клетки.
Опыление
Оплодотворению у цветковых растений предшествует опыление — процесс переноса пыльцы с пыльников на рыльце пестика. При всем разнообразии видов опыления можно выделить два основных его типа: самоопыление и перекрестное опыление. При самоопылении пыльца попадает с тычинки на рыльце пестика в пределах одного цветка (фиалка, ячмень, арахис, овес, просо и др.). Самоопыление может происходить как в открытых цветках, так и в закрытых. Оно имеет важное биологическое значение, поскольку в таком случае оплодотворение гарантировано, и, к тому же, формируются генетически однородные популяции растений, которые могут успешно расселяться. Самоопыление широко используется в селекции культурных растений. Однако самоопыление часто ограничивает приспособляемость растений к условиям окружающей среды и поэтому не способствует процветанию вида. В связи с этим у многих растений существуют препятствия для самоопыления, которые заключаются в разновременности созревания тычинок и пестиков, их различной длине и наличии механизмов самонесовместимости, когда собственная пыльца не прорастает на рыльце пестика. Однако самым надежным механизмом ограничения самоопыления является двудомность.
При перекрестном опылении новый организм обогащается наследственной информацией отцовского организма, что, безусловно, благоприятно сказывается на его приспособляемости к условиям окружающей среды. Перекрестное опыление может осуществляться как с помощью факторов неживой природы, так и живыми организмами.
Перенос пыльцы происходит при помощи ветра (анемогамия, анемофилия) и воды (гидрогамия, гидрофилия). Ветроопыляемых растений среди покрытосеменных немало. Ветром опыляются почти все злаки, осоковые, сережкоцветные и др. У ветроопыляемых растений цветки небольшие, с невзрачным околоцветником или голые, пыльники и рыльца пестика часто свешиваются из цветка, образуют большое количество легкой и сухой пыльцы, которая переносится на расстояние до 5 км. Так, в одной сережке орешника может быть около 4 млн пылинок, а в соцветии кукурузы — еще больше. Многие ветроопыляемые растения цветут вообще до появления листьев, как, например, береза и орешник. Гидрофилия в основном присуща растениям, целиком погруженным в воду (роголистник, стрелолист и др.). Их цветки имеют длинные нитевидные пыльники, а их пыльца лишена оболочки, защищающей от высыхания.

Опыление при помощи животных в основном производится муравьями (мирмекофилия, мирмекогамия), другими насекомыми (энтомофилия, энтомогамия), птицами (орнитофилия, орнитогамия) и т. д. Однако в основном пыльцу переносят все же насекомые (до 70 % покрытосеменных растений). Отличительным признаком цветков, опыляемых насекомыми и птицами, являются ярко окрашенные околоцветники, благодаря которым они еще издали заметны для опылителей, но и зачастую выделяют нектар, привлекающий их. У многих растений этой группы цветки собраны в соцветия. Приспособление к опылению насекомыми зашло у многих растений настолько далеко, что они не способны даже плодоносить без опыления. Например, в Австралии, куда завезли клевер, не было таких насекомых, которые могли бы опылять его, и только завоз шмелей из Европы способствовал выращиванию этой культуры. Пчелы превращают собранный нектар в запасное вещество — мед, который используется в качестве лечебного средства. Чтобы собрать 1 г меда, пчела должна посетить свыше 7 млн цветков. Важными медоносными растениями в Украине являются белая акация, липа, гречиха. Опыление птицами свойственно растениям тропиков. Это большей частью ярко окрашенные орхидеи, канны, фуксии и др., образующие много нектара. К ним за нектаром прилетают колибри, нектарницы, медососы и другие птицы, которые не садятся на цветок, а парят возле него, высасывая нектар. При этом пыльца приклеивается к их головкам.
Кроме естественных способов опыления существует также и искусственное опыление, которое применяется человеком для выведения новых сортов культурных растений и повышения урожайности перекрестноопыляемых растений. Например, ранее в посевах подсолнечника ходили люди в специальных рукавицах, которыми слегка хлопали по раскрывшимся корзинкам. Особую роль искусственное опыление играет при неблагоприятных погодных условиях (пониженные температуры, высокая влажность), когда естественное опыление насекомыми или ветром затруднено.
Двойное оплодотворение
Оплодотворение у покрытосеменных отличается от всех остальных растений, так как оно является двойным. Этот процесс был открыт в 1898 году русским ученым С. Г. Навашиным, работавшим в Киевском императорском университете Св. Владимира.
После попадания на рыльце пестика пыльца прорастает благодаря образованию вегетативной клеткой пылинки пыльцевой трубки. Она проникает через рыльце пестика до пыльцевхода в семязачатке. По пыльцевой трубке двигаются два спермия. Когда пыльцевая трубка достигает семязачатка, она лопается, и спермии оказываются вблизи яйцеклетки и центральной клетки, после чего сливаются с ними. В результате двойного оплодотворения из яйцеклетки и первого спермия образуется зигота, а из центральной клетки и второго спермия — триплоидная клетка. Двойноеоплодотворение играет важную роль у цветковых, поскольку обеспечивает высокую приспособляемость к условиям окружающей среды.
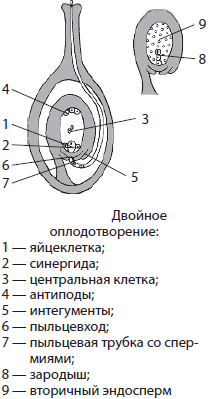
В конечном итоге из семязачатка формируется семя: интегументы дают начало семенной кожуре, зигота — в результате многократных делений — зародышу, а триплоидная клетка — вторичному эндосперму. Последний является запасом питательных веществ для последующего развития зародыша. Запасные вещества могут оставаться в семенах в эндосперме, но могут также переходить в сам зародыш и откладываться в его листочках — семядолях. В некоторых случаях питательные вещества полностью расходуются в процессе образования семени и поэтому семя лишено их, как, например, у многих орхидей. В таких случаях для прорастания семени необходимо наличие гиф симбиотического гриба.
Превращение семязачатка в семя происходит на материнском растении за счет его питательных веществ. Созревшие семена, содержащие в себе зародыши и запасы питательных веществ для их развития, могут долгое время находиться в состоянии покоя, то есть не прорастать в течение ряда лет. Рекорд хранения семян принадлежит семенам лотоса, пролежавшим на дне болота более 10 тыс. лет, а также еще более древним семенам люпина, найденным в Канаде. Это дает им существенное преимущество перед спорами, которые должны прорастать сразу после высыпания, и гибнут, если условия для их развития неблагоприятны. Покой семян может быть глубоким или неглубоким.
Однако есть некоторые растения, у которых семена прорастают прямо на материнском растении, что зачастую имеет приспособительное значение. Это происходит, например, у растения мангровых зарослей — ризофоры, вследствие чего опадающий плод втыкается корнем в ил под деревом и закрепляется, не будучи подхваченным приливом. Но у культурных растений такое прорастание считается существенным недостатком, так как портит урожай. Ранее такое случалось с рожью в дождливые и теплые годы, что грозило голодом. В наше время это бывает у некоторых сортов кукурузы.
Основные отделы растений
Низшие растения
Низшие растения являются наиболее древней группой растений на Земле, объединяющей как одно-, так и многоклеточные водоросли. Слово «водоросль» подразумевает, что растение растет в воде, однако не всякое водное растение является водорослью. Например, хорошо известные кувшинки, роголистник и другие — это высшие растения, освоившие водную среду. Водоросли обитают не только в воде, но и в почве, на коре деревьев, во льдах и т. д., а также входят в состав лишайников. Водоросли водной среды обитания по способу жизни делятся на планктонные и бентосные. Если к планктону относят живые существа, обитающие в толще воды, то бентос представляет собой совокупность живых организмов, живущих на дне водоемов или на значительной глубине.
Слоевище водорослей крайне разнообразно по строению, форме, величине, окраске и т. д. Именно окраска слоевища зачастую отражается в названиях различных таксонов (групп) водорослей: зеленые, бурые, красные и т. д. Окраска водорослей обусловлена пигментами (красящими веществами). Все водоросли содержат хлорофилл, но у многих он маскируется другими пигментами, например, каротиноидами или фикобилинами, которые и придают телу водоросли специфическую окраску. Хлорофилл находится у водорослей в хлоропластах, которые отличаются как по форме, так и количеством. Клеточная оболочка у них целлюлозная, но у некоторых представителей покрывается слизью или минерализуется, то есть в ней откладываются минеральные соли. По строению таллом бывает одноклеточным, колониальным или многоклеточным. Многоклеточные талломы бывают нитчатыми, кустистыми и пластинчатыми.
Размножаются водоросли бесполым, вегетативным и половым способами. При бесполом размножении путем спорообразования содержимое одной клетки многократно делится и образуется значительное количество одноклеточных подвижных спор, которые называются зооспорами. Каждая из них дает начало новой особи. У некоторых неподвижных водорослей бесполое размножение осуществляется не зооспорами, а неподвижными спорами, лишенными жгутиков. Споры образуются в клетках, не отличающихся по форме от других клеток, или в особых клетках-спорангиях, которые могут иметь иную форму и величину, чем вегетативные.
При вегетативном размножении новые особи возникают из обрывков нитей и кусков слоевищ многоклеточных водорослей, а также при распаде колоний и делении одноклеточных представителей надвое. Иногда для такого размножения существуют специальные образования.
Половое размножение в классическом виде заключается в слиянии двух половых клеток, в результате чего образуется зигота, прорастающая затем в новую особь, однако у водорослей могут сливаться как половые, так и вегетативные клетки или даже одноклеточные особи целиком. Для некоторых водорослей характерен особый половой процесс — конъюгация.
Следует отметить, что у одних водорослей споры и гаметы образуются на одном растении, тогда как у других органы бесполого и полового размножения развиваются у особей разных поколений.
Водоросли не являются систематической группой, так как эти организмы имеют различное строение и происхождение, тем не менее, по традиции, к ним относят до 13 отделов водных растений, наиболее изученными из которых являются красные, бурые, зеленые и диатомовые водоросли.
Отдел Красные водоросли, или Багрянки
Отдел Красные водоросли включает около 5 000 видов в основном морских многоклеточных водорослей, хотя в настоящее время известны уже пресноводные и почвенные виды. Окраску их таллому от голубовато-стального до малиново-красного и даже черного придают хлорофилл и дополнительные пигменты — фикобилины. Красные водоросли — наиболее глубоководные, так как они были обнаружены на глубине 268 м в районе Багамских островов, где освещенность составляет около 0,0005 % ее значения у поверхности моря. Основное запасное вещество красных водорослей — багрянковый крахмал. Размножаются багрянки бесполым, вегетативным и половым способами, при этом даже споры и половые клетки лишены жгутиков, что коренным образом отличает их от других водорослей.
Основными представителями красных водорослей являются порфира, родимения, калитамнион, немалион, кораллина, анфельция и церамия, обитающие в морях, тогда как в пресных водоемах в нашей стране можно встретить, например, батрахоспермум.
Красные водоросли играют важную роль в морских экосистемах, так как служат кормом для многих животных и дают им приют, обогащают воду кислородом, поглощают углекислый газ. Некоторые красные водоросли используют в пищу, на корм скоту, в качестве удобрения. Также из них получают йод, бром, заменители крови и лекарственные препараты, препятствующие свертыванию крови. Огромное значение имеют родимения и порфира, служащие источниками агарагара, используемого в микробиологической промышленности и биотехнологии для изготовления питательных сред, а также в пищевой, бумажной и текстильной промышленностях. В Японии, Украине и других странах порфиру даже культивируют.
Отдел Бурые водоросли
Отдел Бурые водоросли объединяет около 1 500 видов исключительно многоклеточных морских организмов. Окраска слоевища бурых водорослей варьирует от зеленовато-оливковой до темно- бурой, будучи обусловленной хлорофиллом и каротиноидами. Размеры тела бурых водорослей могут достигать 60 и более метров (макроцистис грушеносная). К субстрату бурые водоросли прикрепляются с помощью специальных выростов — ризоидов, но впоследствии могут отрываться от него и новых ризоидов не образовывать. У них имеются достаточно хорошо сформированные ткани, а органы полового и бесполого размножения, в отличие от всех остальных водорослей, могут быть многоклеточными. Запасное вещество бурых водорослей — растворимый углевод ламинарин, накапливающийся в цитоплазме. Размножаются бурые водоросли вегетативно, бесполым или половым способом. Для них характерно чередование бесполого и полового поколений с преобладанием первого в жизненном цикле.
К данному отделу принадлежат ламинария, фукус пузырчатый , саргассум и макроцистис.
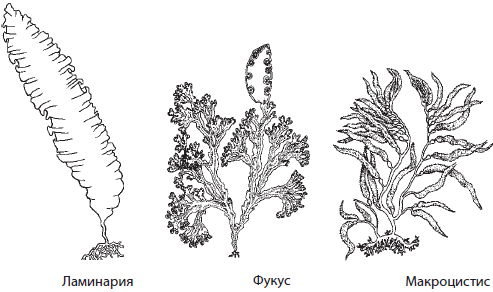
Ламинария, или «морская капуста» — бурая водоросль с талломом, на котором пластинчатые «листья» прикрепляются к простому или разветвленному «стволу». Длина некоторых ламинарий может достигать до 20 м. Она накапливает значительное количество питательных веществ и йода, поэтому во многих странах ее употребляют в пищу и даже разводят искусственно.
Фукус пузырчатый — крупная бурая водоросль до 70 см в длину, заросли которой покрывают сплошным ковром дно в прибрежной зоне северных морей (Белого, Баренцева, Охотского). На концах слоевища заметны своеобразные вздутия, которые помогают ему удерживаться на плаву. Эти вздутия лопаются под ногами, если идти по ним во время отлива.
Саргассум — род многолетних морских водорослей длиной до 1 м, которые, отрываясь от субстрата в Мексиканском заливе, образуют сплошные скопления в Саргассовом море, названном в их честь.
Бурые водоросли играют крайне важную роль в морских экосистемах, так как являются важнейшим поставщиком органического вещества и дают приют настолько многим видам организмов, что великий английский биолог Ч. Дарвин сравнивал подводные леса Южного полушария с наземными лесами тропических областей и считал, что уничтожение леса в какой-либо стране привело бы к гибели значительно меньшего числа видов животных, чем уничтожение водорослей. Бурые водоросли широко используются в пищу, для изготовления лекарственных препаратов, получения йода и т. д.
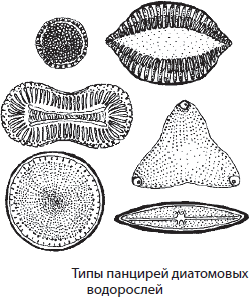
Отдел Диатомовые водоросли
Отдел Диатомовые водоросли объединяет около 20 тыс. видов одноклеточных и колониальных организмов, имеющих кремнеземный панцирь. Размеры тела диатомовых водорослей крайне незначительны — в среднем около 0,02–0,05 мм. Панцирь имеет две створки: верхнюю и нижнюю. Окраска их обычно грязно-желтая, что обусловлено наличием дополнительных пигментов фотосинтеза — каротиноидов, маскирующих хлорофилл. Основным запасным веществом диатомовых водорослей являются полисахариды хризоламинарин и волютин, реже — масло. Размножение осуществляется бесполым и половым способами. Диатомовые водоросли преобладают в морских и пресноводных экосистемах, обитают в почве.
Характерными представителями диатомовых водорослей являются пиннулярия, навикула, цимбелла, мелозира и др.
Диатомовые водоросли играют важную роль в водных экосистемах, поскольку служат основным кормом для значительного количества водных организмов, в том числе молодняка многих рыб. По питательной ценности они не уступают пищевым растениям. Отмирая, эти водоросли опускаются на дно и служат пищей для бактерий и простейших. Они также имеют исключительное значение в осадконакоплении, образуя диатомовые илы. Так, осадочная порода диатомит состоит на 50–80 % из их панцирей, что способствует ее использованию в качестве абразивного и поглощающего материала, а также фильтра. Панцири диатомовых водорослей также используются в геологии и палеонтологии в качестве «руководящих ископаемых», по которым можно датировать возраст горных пород и останков, найденных в них.
Отдел Зеленые водоросли
К отделу Зеленые водоросли относят 20–25 тыс. видов водорослей с преимущественно зеленой окраской таллома вследствие преобладания хлорофилла. Они представлены одноклеточными, колониальными и многоклеточными формами.
Многоклеточные зеленые водоросли могут иметь нитчатый или пластинчатый таллом. Основным запасным веществом зеленых водорослей является крахмал. Размножение осуществляется бесполым, вегетативным и половым способами. Они заселили все возможные экологические ниши: соленые и пресные водоемы, почву, камни, ледники, гейзеры и пр.
Характерными представителями зеленых водорослей являются хламидомонада, вольвокс, хлорелла, спирогира, улотрикс и ульва. К ним же относят и эвглену зеленую.
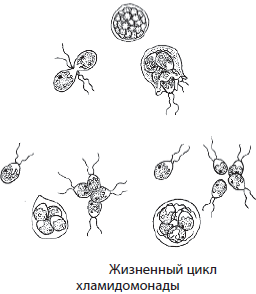
Хламидомонада — одноклеточная зеленая водоросль, обитающая во всех пресных водоемах, лужах и других временных водоемах.
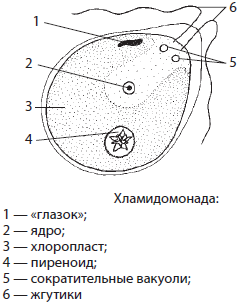
Ее клетка имеет грушевидную форму и снабжена двумя жгутиками. Единственный хлоропласт хламидомонады имеет чашевидную форму. В передней части клетки находится красный «глазок», который ориентирует хламидомонаду на свет. Имеются также две сократительные вакуоли, которые удаляют из клетки излишнюю воду. При неблагоприятных условиях хламидомонады теряют жгутики, покрываются слизистыми капсулами и переходят в состояние покоя. Если такую клетку поместить в воду, она возобновит жизнедеятельность. Размножаются хламидомонады бесполым и половым способами. При бесполом размножении в результате двух митотических делений клетки образуются четыре подвижные зооспоры, служащие для расселения. Половое размножение сопровождается формированием в материнских клетках похожих на зооспоры гамет со жгутиками, которые затем сливаются. Из зиготы хламидомонад в результате мейоза образуется сразу четыре гаплоидных дочерних особи. Хламидомонады широко используются в лабораторных исследованиях.
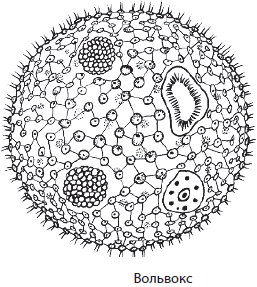
Вольвокс — колониальная зеленая водоросль сферической формы. Ее клетки расположены в один слой, а внутреннее пространство колонии заполнено слизью. Клетки вольвокса соединены между собой цитоплазматическими мостиками и делятся на две группы: вегетативные и генеративные. Вегетативные клетки похожи по строению на клетки хламидомонады, они осуществляют процесс фотосинтеза, но не могут принимать участия в размножении, тогда как генеративные клетки выполняют только эту функцию. Вольвоксы размножаются вегетативным и половым способами. При вегетативном размножении генеративная клетка делится несколько раз с образованием дочерней колонии, которая затем выпадает вовнутрь материнской и освобождается только после ее гибели. При половом размножении в других генеративных клетках формируются гаметы, сливающиеся с образованием зиготы. В зиготе образуется одна зооспора, которая после многократных делений превращается в новую колонию. Обитает вольвокс в пресных водоемах.
Хлорелла — одноклеточная пресноводная водоросль, клетка которой имеет сферическую форму. Хлореллы встречаются в пресных и морских водоемах, на коре деревьев, в почве и других влажных местах обитания. Размножение хлорелл осуществляется бесполым способом. Благодаря высокой эффективности использования солнечного света (до 12 %), накоплению белка и быстрому росту культуры они используются в лабораторных исследованиях и для обеспечения жизнедеятельности экипажей космических станций и подводных лодок.
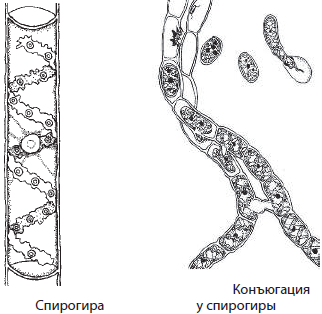
Спирогира — многоклеточная нитчатая пресноводная водоросль, спирально уложенные хлоропласты которой имеют лентовидную форму. Размножается спирогира вегетативно и половым способом. Вегетативное размножение осуществляется путем разрыва нитей; половой процесс называется конъюгацией. При конъюгации две нити спирогиры подходят друг к другу, их клетки формируют конъюгационные мостики, по которым содержимое клеток одной нити перетекает в другую и сливается с ее клетками с образованием зиготы. Прорастание зиготы сопровождается последовательными делениями, в результате которых развивается новая особь.
Улотрикс — небольшая (до 10 см) многоклеточная нитчатая водоросль, образующая сплошные дерновины на подводных предметах в реках и ручьях. Неразветвленные слоевища улотрикса прикрепляются к субстрату с помощью вытянутой клетки — ризоида. Хлоропласты водоросли имеют подкововидную форму. Улотрикс размножается вегетативно, бесполым или половым способами. При вегетативном размножении нить улотрикса распадается на несколько частей, каждая из которых дает начало новому организму. Бесполое размножение осуществляется с помощью зооспор. При половом размножении в клетках слоевища образуются гаметы, снабженные жгутиками.
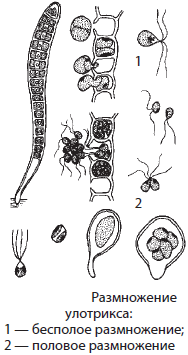
Они сливаются с другими гаметами собственного таллома или других нитей и образуют зиготу, которая вскоре оседает на дно и переходит в состояние покоя. Прорастание зиготы сопровождается ее делением с образованием 4–8 зооспор, дающих начало новым особям.
Ульва — многоклеточная пластинчатая морская водоросль, в жизненном цикле которой происходит чередование полового и бесполого поколений. Ульву, или «морской салат», человек активно употребляет в пищу.
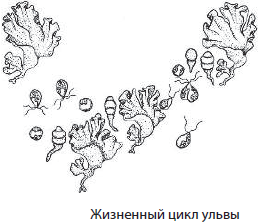
Значение зеленых водорослей в природе и жизни человека трудно переоценить, поскольку они обеспечивают приток органического вещества в водных и других экосистемах, образуют значительное количество кислорода, поступающего в атмосферу, принимают участие в круговороте веществ в природе и т. д. Некоторые зеленые водоросли употребляет в пищу человек, их используют на корм скоту, как удобрение, как индикаторы загрязнения окружающей среды и т. д. Вместе с тем они могут вызывать «цветение» воды, приводящее к замору рыбы и других водных животных.
Высшие споровые растения
Выход растений на сушу более 400 млн лет назад был сопряжен с формированием тканей и органов, строение которых постепенно усложнялось в процессе эволюции. Прогрессивному усложнению морфологии и анатомии растений соответствовали изменения и в их онтогенезе: в жизненном цикле наземных растений происходит правильное чередование бесполого и полового размножений и связанное с этим чередование бесполого и полового поколений. Бесполое поколение называется спорофитом, а половое — гаметофитом. На спорофите (2n) развиваются спорангии, в которых в результате мейоза формируются споры (n). Споры являются одноклеточными образованиями с небольшим запасом питательных веществ и довольно прочной защитной оболочкой. Из споры вырастает гаметофит (n). Именно наличие отдельной фазы гаметофита в жизненном цикле является характерным признаком высших споровых растений. На гаметофите развиваются генеративные органы — гаметангии, которые подразделяются на женские — архегонии — и мужские — антеридии. В архегониях образуются одиночные яйцеклетки (n), тогда как в антеридиях — множество подвижных сперматозоидов (n). Оплодотворение у высших споровых растений, как и у низших, требует наличия хотя бы капельно-жидкой влаги. В результате оплодотворения образуется зигота (2n), из которой сначала формируется многоклеточный зародыш, развивающийся впоследствии в новый спорофит. Современные высшие споровые растения представлены отделами Моховидные, Плауновидные, Хвощевидные и Папоротниковидные.
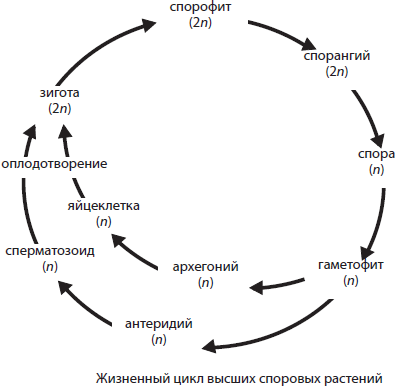
Следует отметить, что у наиболее высокоорганизованных представителей плаунов и папоротников на спорофитах развиваются два типа спорангиев: мегаспорангии и микроспорангии. В мегаспорангиях формируются мегаспоры, вырастающие затем в женские гаметофиты с архегониями, тогда как в микроспорангиях образуются микроспоры, дающие начало мужским гаметофитам с антеридиями. Дальнейшие события в жизненных циклах таких представителей не отличаются от вышеописанных. Растения, образующие два типа спор, называются разноспоровыми, а все остальные — равноспоровыми. Именно разноспоровые растения, скорее всего папоротники, стали предками семенных растений.
Отдел Моховидные
Современные моховидные представлены примерно 25 тыс. видов споровых растений, в жизненном цикле которых преобладает гаметофит. Размеры мхов незначительны: большинство их не достигает в высоту и 10 см, хотя имеются и 40-сантиметровые виды. Мхи обитают в основном во влажных местах. Большое видовое разнообразие мхов характерно для северных широт, в том числе зоны тундры, однако и в тропических областях встречается немало видов этих организмов. Гаметофит моховидных может иметь листостебельное или слоевищное строение, но, в отличие от низших растений, у них имеются ткани (покровная и основная).
Наибольшее число представителей отдела насчитывают два класса: Печеночники и Листостебельные мхи. Характерными представителями печеночников являются маршанция и риччия, а листостебельных — политрихум и сфагнум.
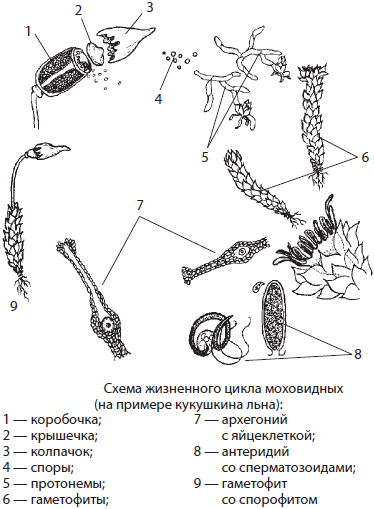
Характерным представителем листостебельных мхов является политрихум, или кукушкин лен, зачастую образующий сплошной покров на сырой почве в лесах, на лугах и болотах. Гаметофит кукушкина льна достигает в высоту 10–20 см. Надземная часть его стебля усеяна жесткими листочками, а от подземной отходят ризоиды, которые служат для прикрепления к субстрату и поглощения воды. Корней у мхов нет.
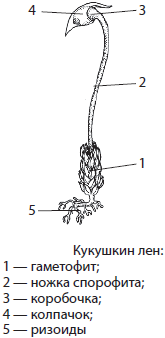
Гаметофиты кукушкина льна раздельнополые. На верхушках женских гаметофитов развиваются архегонии с яйцеклетками, а на верхушках мужских — антеридии со сперматозоидами. Оплодотворение у политрихума происходит в дождливую погоду или при сильной росе. Из зиготы прямо на материнском растении вырастает спорофит. Зрелый спорофит кукушкина льна — спорогон — состоит из ножки и коробочки с крышечкой и колпачком. Он паразитирует на гаметофите, поглощая из его тканей воду и растворенные в ней питательные вещества с помощью специального выроста ножки — гаустории. В коробочке спорофита формируются споры, которые после созревания высыпаются и разлетаются на значительное расстояние, осуществляя бесполое размножение. Из спор прорастают протонемы, или предростки, дающие начало новым гаметофитам.
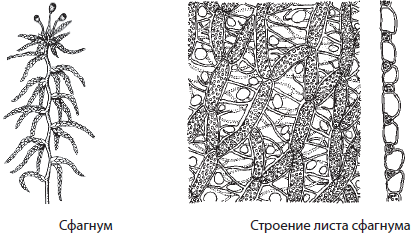
Сфагнум — белый болотный мох, образующий часто сплошной покров в избыточно увлажненных местах. Цвет растений сфагнума обусловлен тем, что часть его клеток специализируется на накоплении воды. За год прирост сфагнума составляет около 3–5 см, и столько же отмирает в его нижней части, однако не разлагается из-за особенностей среды обитания сфагнума. В отличие от кукушкина льна, гаметофиты сфагнума однодомные, а коробочки сфагнума имеют округлую форму. Сфагнум играет первостепенную роль в процессах торфообразования.
Маршанция имеет слоевищный вильчато ветвящийся гаметофит, на нижней стороне которого развиваются ризоиды. Вегетативное размножение маршанции осуществляется с помощью выводковых корзинок, а половое — с помощью специальных выростов на ножках. Маршанции растут во влажных местах, где отсутствует травяной покров.
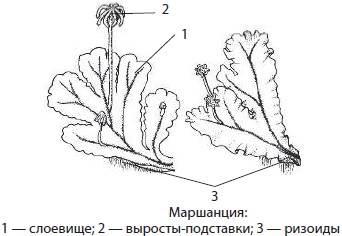
Риччия, или водяной мох, освоила водоемы, например, на Дальнем Востоке, и достаточно популярна у аквариумистов, поскольку в ней могут укрываться мелкие рыбки.
Значение мхов в природе велико — они защищают почвы от высыхания и эрозии, участвуют в образовании торфа, широко используются в химической промышленности для получения различных органических веществ. Ранее сфагнум применялся как заменитель ваты, так как он гигроскопичен и обладает антисептическими свойствами. С другой стороны, мхи также способствуют быстрому заболачиванию почв из-за образования сплошного покрова, они также вытесняют травы. Из-за накопления в мхах различных несъедобных веществ их слабо поедают животные. Торф образуется на болотах очень медленно — его прирост составляет 1 см за 10 лет, поэтому бесконтрольное изъятие мхов во многих местах привело к нарушению экологического равновесия и потребовало срочных мер по защите реликтовых болот.Отдел Плауновидные
К плауновидным относят древнейшие из споровых растений, в жизненном цикле которых преобладает спорофит. Ныне встречается около 1000 видов плауновидных. Несмотря на то, что ранее среди плаунов встречались и древесные формы, до наших дней сохранились в основном многолетние травянистые растения.
В наших хвойных лесах часто встречается плаун булавовидный с вечнозелеными стелющимися и поднимающимися побегами, густо усаженными мелкими шиловидными листочками. На стебле плауна образуются тонкие придаточные корни. Для представителей этого отдела высших споровых уже характерно наличие проводящих, механических и иных тканей.
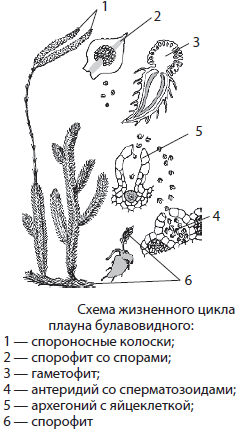
Споры у плауна созревают на особых листьях — спорофиллах, собранных в спороносные колоски на верхушках побегов. После высыпания из спор вырастают гаметофиты, ведущие подземный образ жизни и фактически паразитирующие на гифах гриба в течение 6–20 лет, до образования на них архегониев и антеридиев. После оплодотворения, для которого необходима влага, из зиготы вырастает спорофит.
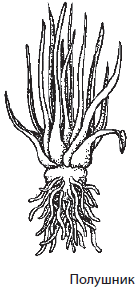
В нашей стране, кроме плауна булавовидного, встречаются и другие представители отдела, например плаун годичный, баранец обыкновенный и полушник озерный. Полушники интересны тем, что являются разноспоровыми растениями и, в отличие от плаунов, освоили водную среду обитания.
В прошлом плауновидные были широко распространены и играли важную роль в наземных экосистемах. Их останки впоследствии образовали залежи каменного угля и нефти, которые широко используются в народном хозяйстве не только как топливо, но и в качестве сырья для химической промышленности. В настоящее время значение плауновидных существенно уменьшилось, так как их не едят животные, а использование ограничивается применением спор в качестве детской присыпки и обсыпки форм для литья в промышленности. Кроме того, их применяют в народной медицине как средство для лечения алкоголизма и для украшения корзинок с крашенками к Пасхе. Бесконтрольный сбор спорофитов и спор этих растений в прошлом привели к тому, что некоторые плауны стали редки и занесены в Красную книгу.
Отдел Хвощевидные
К хвощевидным относят около 30 видов ныне живущих многолетних травянистых растений, хотя ранее они были представлены в основном древесными и кустарниковыми формами. Хвощи преимущественно распространены во влажных местах. Характерным представителем отдела является хвощ полевой. Побеги хвоща расчленены на узлы и междоузлия. В узлах расположены мутовки побегов и крошечные чешуевидные листочки. У хвощей функцию фотосинтеза выполняют только стебли, тогда как листья даже не содержат хлорофилла. Стебель хвоща имеет многочисленные полости, заполненные водой или воздухом. Клетки кожицы стебля пропитаны кремнеземом, что делает его очень жестким и несъедобным для животных. Подземная часть побега хвоща представлена корневищем, в котором накапливаются питательные вещества. На корневище ежегодно образуются надземные побеги и придаточные корни.

Споры хвощевидных образуются в специальных спороносных колосках, расположенных на верхушках побегов. Эти колоски могут развиваться как на обычных, вегетативных побегах, так и на специальных — генеративных, которые появляются весной, раньше вегетативных. После созревания споры хвоща высыпаются, и при благоприятных условиях из них вырастают зеленые пластинчатые гаметофиты. На гаметофите развиваются архегонии и антеридии, в которых созревают соответствующие половые клетки. Оплодотворение у хвощей, как и у других высших споровых растений, зависит от воды. После слияния сперматозоида с яйцеклеткой из зиготы у них сначала развивается зародыш, питающийся за счет гаметофита до тех пор, пока не сформирует собственные корни и надземный побег, а он, в свою очередь, дает начало взрослому спорофиту.

Помимо хвоща полевого, в России встречается еще несколько видов хвощей: хвощ речной, хвощ лесной и другие.
Значение хвощей в настоящее время в природе невелико, так как они несъедобны для животных, однако их предки сыграли немаловажную роль в образовании залежей каменного угля. Хвощи применяют в качестве абразивного средства для чистки посуды и шлифовки различных изделий, в медицине — как мочегонные и кровоостанавливающие средства. Молодые побеги хвощей ранее использовали в пищу. Многие хвощи являются индикаторами кислых почв. Среди них имеются ядовитые виды и злостные сорняки.
Отдел Папоротниковидные
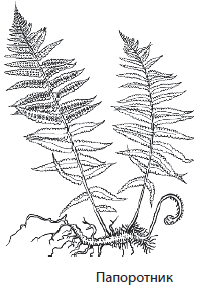
К папоротниковидным относят около 12 000 видов споровых растений, долгоживущий спорофит которых имеет крупные, часто сильно рассеченные черешковые листья — вайи. На нижней стороне вай размещаются отдельные спорангии или их группы — сорусы. Папоротники распространены повсеместно, особенно в тропиках, ряд представителей отдела освоили водную среду (марсилия, сальвиния, азолла).
Спорофит папоротников умеренного климата имеет корневище с придаточными корнями и хорошо развитые листья, тогда как в южных областях планеты встречаются древовидные и лиановидные формы этих растений. Вайи папоротников могут достигать в длину от 2–4 мм до 6 м, среди них встречаются как простые, так и перисто- рассеченные. Особенностью листьев папоротников является длительное нарастание верхушкой, вследствие чего молодой лист свернут улиткой и постепенно раскручивается.
На нижней стороне зрелых листьев могут образовываться многочисленные спорангии, собранные в группы — сорусы, иногда даже прикрытые специальным покрывальцем — индузием. Созревшие споры рассеиваются с помощью ветра, из них вырастают небольшие сердцевидные гаметофиты до 1 см в диаметре. На нижней стороне гаметофита развиваются антеридии, архегонии и ризоиды. В архегониях образуются женские половые клетки — яйцеклетки, а в антеридиях — многожгутиковые сперматозоиды. Для оплодотворения папоротникам, как и остальным высшим споровым растениям, обязательно необходимо наличие капельно- жидкой влаги. Из зиготы вырастает спорофит, который первое время паразитирует на гаметофите.
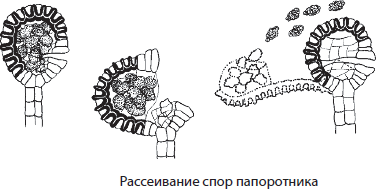
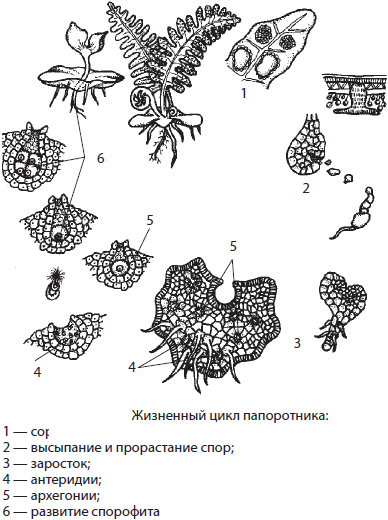
Большинство папоротников — равноспоровые растения, однако часть представителей отдела, освоивших водную среду, относятся к разноспоровым, как марсилия, сальвиния и азолла.
В России произрастает около 100 видов папоротников, в том числе щитовник мужской, кочедыжник женский, орляк, страусник обыкновенный и др.
Папоротники сыграли значительную роль в образовании залежей каменного угля, а в настоящее время широко используются в пищу как лекарственные растения (глистогонное, противовоспалительное средства), а также культивируются на рисовых полях для обогащения их азотом. Представители отдела популярны в качестве декоративных растений: адиантум венерин волос, платицериум («олений рог»), нефролепис и др. Некоторые папоротники (марсилии) культивируются в качестве аквариумных растений.
Семенные растения
Общая характеристика
Семенные растения являются господствующей в настоящее время группой растений. Их широкое распространение в значительной степени обусловлено возникновением семени, а также независимостью полового процесса даже от капельно-жидкой влаги.
В жизненном цикле семенных растений преобладает спорофит, тогда как гаметофиты настолько редуцированы, что развиваются из спор непосредственно на спорофите. Все семенные относятся к разноспоровым растениям, так как на их спорофитах развиваются мега- и микроспорангии с мега- и микроспорами соответственно. Мегаспорангий семенных видоизменен в семязачаток с мегаспорами, одна из которых развивается в женский гаметофит. На последнем впоследствии формируются яйцеклетки.
Микроспоры прорастают в редуцированные мужские гаметофиты, не покидая при этом оболочки споры. Мужской гаметофит семенных состоит обыкновенно из двух клеток — вегетативной и генеративной и называется пыльцевым зерном, или пылинкой.
Оплодотворению у семенных предшествует опыление — перенос пыльцевого зерна с помощью ветра или иных факторов к женскому гаметофиту. Здесь вегетативная клетка мужского гаметофита прорастает в пыльцевую трубку, по которой движутся образовавшиеся из генеративной клетки спермии вплоть до слияния с яйцеклеткой. Именно появление пыльцевой трубки позволило семенным растениям преодолеть зависимость от воды в процессе оплодотворения.
В результате оплодотворения из семязачатка формируется семя — многоклеточное образование, содержащее зародыш — маленький спорофит, запас питательных веществ и достаточно прочные покровы, что обеспечивает ему возможность сохраняться длительное время до наступления благоприятных условий. К тому же семя зачастую снабжено специальным приспособлением для распространения (прицепками, крючками и др.).
В целом семенные характеризуются более совершенным строением тканей и органов.
К семенным растениям в настоящее время относят два отдела высших растений: Голосеменные и Покрытосеменные, или Цветковые.
Отдел Голосеменные
К голосеменным относят около 800 видов семенных растений, не формирующих цветков и плодов. Их семязачатки лежат открыто (голо) на чешуйках женских шишек (отсюда название отдела). Почти все голосеменные — древесные, исключительно наземные растения, многие из которых образуют леса в различных частях света. Для них характерно более совершенное строение тканей и органов по сравнению с высшими споровыми растениями, в частности, у них появляется главный корень, который развивается из зародышевого корешка, а рост стебля и корня в толщину обеспечивается делением клеток камбия, который год за годом откладывает все новые годичные кольца древесины. Стебли и корни также могут покрываться пробкой и коркой, что не присуще высшим споровым.
Характерным представителем голосеменных является сосна обыкновенная. Как правило, это стройное дерево высотой до 30–40 м. На удлиненных побегах сосны расположены укороченные, несущие пучки из двух листьев-хвоинок. Хвоинки опадают через 2–3 года, поэтому сосна относится к вечнозеленым растениям. Ткани сосны, в частности мякоть листа, первичная кора, луб и древесина пронизаны смоляными ходами с живицей, выполняющей защитную функцию. Хвоинки также выделяют летучие бактерицидные вещества — фитонциды.
Как и все голосеменные, сосна — разноспоровое растение. На нижней стороне чешуек мужских шишек, лежащих в основании развивающихся побегов, весной образуются микроспорангии — пыльцевые мешки — с микроспорами — пылинками. Не покидая оболочки пыльцевого зерна, из микроспор вырастают сильно редуцированные мужские гаметофиты, состоящие из двух клеток — вегетативной и генеративной. Пылинка зачастую имеет два воздушных мешка для облегчения его переноса ветром.
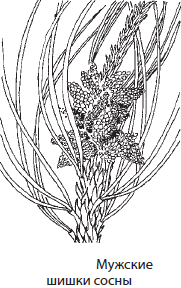
На чешуйках ярко окрашенных женских шишек, находящихся на концах развивающихся побегов, развивается по два видоизмененных мегаспорангия — семязачатка. Семязачатки покрыты интегументом и содержат материнскую ткань — нуцеллус. В процессе спорообразования в семязачатке остается по одной мегаспоре, которая прорастает в многоклеточный женский гаметофит (n). На гаметофите образуется два архегония, в каждом из которых формируется по яйцеклетке.
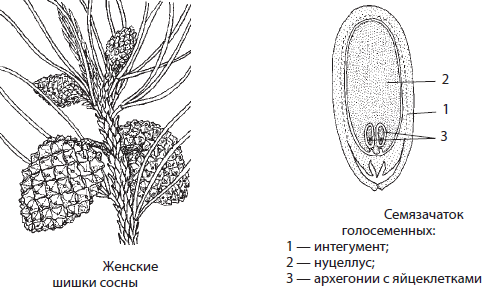
К моменту созревания пыльцы семязачатки выделяют небольшую каплю клейкой жидкости, к которой прилипают пылинки. Затем семязачаток поглощает жидкость и втягивает пылинки внутрь шишки. Когда пылинка попадает на женскую шишку, она образует пыльцевую трубку, по которой начинают двигаться две мужские половые клетки — спермия, образовавшиеся в результате деления генеративной клетки. Рост трубки вскоре приостанавливается, а сама шишка зеленеет, деревенеет и разрастается. Через год после опыления в семязачатках созревают яйцеклетки, а пыльцевая трубка дорастает до них. Один спермий оплодотворяет яйцеклетку, а оставшиеся спермий и яйцеклетка погибают. Из зиготы формируется зародыш (2n) с несколькими семядолями, а из клеток женского гаметофита — первичный эндосперм (n), тогда как интегумент превращается в семенную кожуру (2n) с крыловидным выростом. Приблизительно через полтора года после опыления женская шишка полностью одревесневает и в хорошую погоду открывается, в результате чего семена разлетаются.

К голосеменным относят семь классов, основным из которых является класс Хвойные. Представителями других классов голосеменных являются вельвичия удивительная, гинкго двулопастный и саговники.
Класс Хвойные включает около 700 видов голосеменных, имеющих игловидные, чешуевидные или иной формы листья, которые чаще всего называют хвоинками. Они имеют толстый слой кутикулы и углубленные устьица для защиты от испарения. Большинство хвойных — вечнозеленые растения, но встречаются и листопадные, как лиственница. Ежегодно у них образуется одно кольцо ветвей, по количеству которых можно определить возраст растения. Многие хвойные выделяют живицу. К классу Хвойные относятся роды: сосна, ель, пихта, лиственница, секвойя, секвойядендрон, кедр, можжевельник, туя, кипарис и др.
Сосны — стройные высокие деревья с прочной древесиной. Их хвоинки собраны в пучки по 2, 3 или 5 и крепятся на укороченных побегах. Семена сосен зачастую снабжены крылатками, у некоторых образуют «орешки», используемые в пищу. Наиболее часто в России встречаются сосна обыкновенная и сосна сибирская.
Ели — высокие деревья с пирамидальной кроной. Хвоинки ели намного короче, чем у сосны. Они жесткие на ощупь и сидят по одной не на укороченных побегах, а прямо на удлиненных. Хвоинки могут жить более 10 лет, после чего опадают. Ветви елей сохраняются всю жизнь, поэтому еловые леса относят к темным. Шишки елей поникающие, опыление, оплодотворение и созревание семян у них происходят в течение одного года. Корневая система елей поверхностная, поэтому сильный ветер может вывернуть растение вместе с корнем. Наиболее распространенными в России считаются ель обыкновенная и ель сибирская.
Пихты — высокие деревья с более мягкой хвоей, чем у елей, и с распадающимися при созревании шишками. Они также образуют темные хвойные леса. Наиболее распространены пихта обыкновенная и пихта одноцветная.
Кедр — род вечнозеленых растений, хвоинки которых собраны в пучки по 30–40, а шишки созревают и рассыпаются на второй-третий год. Известно четыре вида кедров, растущих в том числе на Кавказе.
Лиственницы — высокие деревья с ежегодно опадающей хвоей, собранной по 10–12 штук на укороченных побегах. Их шишки созревают и раскрываются чаще в тот же год, когда произошло опыление, но остаются на ветвях еще в течение нескольких лет. Древесина лиственниц чрезвычайно прочна и не боится влаги, что позволяет широко использовать ее в строительстве, в том числе для изготовления свай. В России произрастают лиственницы сибирская и даурская.
Можжевельник — род невысоких растений, среди которых имеются как невысокие деревья, так и кустарники, в том числе и стелющиеся. Их хвоинки имеют игловидную или чешуевидную формы, а чешуйки шишек срастаются, образуя шишкоягоды. К ним относятся можжевельник обыкновенный, можжевельник казацкий и др.

К роду туя относятся древесные или кустарниковые виды голосеменных с чешуевидными, прижатыми к ветвям, листьями. Их мелкие шишки созревают и раскрываются в тот же год, когда произошло опыление. Представителями рода являются туя западная, туя гигантская и др.
Довольно редкие в наше время тиссы чаще всего принимают форму вечнозеленого приземистого кустарника с двурядно расположенными ланцетными или линейными листьями. Их семена окружены сочным ярко-красным придатком и весьма привлекательны на вид, однако все органы этого растения, в том числе и семена, ядовиты, поэтому следует избегать контакта с ними. В России встречаются в основном тиссы ягодный и остроконечный.
К хвойным относят и такие виды, как секвойи и секвойядендроны, достигающие в высоту нескольких десятков метров и в толщину — более 10 м, живущие несколько тысяч лет.
Интересны также «живое ископаемое» — гинкго двулопастный, или «серебряный абрикос» из Китая и Японии, дающий семена с серебристо- оранжевым придатком и имеющий вееровидные лопастные листья, и вельвичия удивительная из пустыни Намиб, два листа которой функционируют в течение всей ее жизни (до 900 лет). В последнее время большой популярностью в качестве декоративного комнатного растения пользуется и представитель саговников — саговник поникающий (цикас), напоминающий пальму с толстым неветвящимся стволом.
Значение голосеменных в природе и жизни человека трудно переоценить, поскольку они образуют обширнейшие леса на севере нашей страны, в Северной Америке и на островах Тихого океана, и обеспечивают приток органического вещества и обогащение атмосферы кислородом. Древесина голосеменных широко используется в строительстве, для изготовления бумаги; смола и канифоль, полученные из них, широко применяются в химической и лакокрасочной промышленностях; из листьев хвойных получают витамин С, а декоративные формы используют в зеленом строительстве. Семена некоторых хвойных (сосна сибирская, пиния) съедобны.

Онлайн-школа «Турбо»
- Прямая связь с преподавателем
- Письменные дз с проверкой
- Интересные онлайн-занятия
- Душевное комьюнити